

1. 서론
2. 복합과학으로서의 자연지리학
1) 기초학문분야 범례의 발전과 차용
2) 비선형 동역학과 시스템 이론
3) 비선형 동역학과 시스템 이론의 연결
4) 소결
3. 생물지형학의 탄생과 발달
1) 지형학, 토양학, 생태학: 학문사적 흐름의 재발견
2) 각 학문사적 발전 과정이 생물지형학의 태동에 미친 영향
3) 생물지형학의 발달사 조망: 주요 학술행사 소개
4) 자연지리학과 생물지형학의 연결 고리
5) 소결
4. 자연지리학과 진화생물학의 만남
1) 생물지형학의 현재
2) 생물지형학의 미래: 진화생물학을 포용하는 가설의 제안
3) 소결
5. 요약 및 결론
1. 서론
자연지리학은 우리가 사는 대자연을 구성하는 다양한 요소들이 시공간상에서 끊임없이 상호작용하는 과정을 연구하는 종합과학이다. 많은 자연지리학자들은 시스템 접근을 도입하여 지구를 기권(atmosphere), 수권(hydrosphere), 암석권(lithosphere), 생물권(biosphere)으로 구분한다(Christopherson and Birkeland, 2017). 일차적으로는 환원주의적 시각에서 각 권역에 대한 깊은 이해를 도모한 후, 궁극적으로는 이들 사이에서 발생하는 에너지와 물질의 흐름을 파악하여 지구 시스템의 작동 원리를 거시적이고 포괄적인 관점에서 설명하고자 한다(Huggett, 1985; Strahler and Strahler, 2003).
우리나라의 자연지리학계는 오랜 시간 암석권에 큰 관심을 기울여 왔다. 그 결과, 산맥 구조와 하계망 체계의 형성을 아우르는 한반도 지형발달사를 이해하는 데 이론적・방법론적으로 상당한 발전을 이루었다(박수진, 2009; 변종민 ・김종욱, 2011; 성영배・유병용, 2014; 홍성찬, 2016; Kim et al., 2020). 이 과정에서, 기후변화와 해수면 변동의 영향을 깊게 고려함으로써 기권과 수권에 대한 탐색도 꾸준히 진행하였다(이광률, 2023, 2024; 최광용, 2024; 최영은 등, 2023; Lee et al., 2022). 반면, 생물권은 우리나라 자연지리학의 학문사적 흐름에서 상당히 작은 부분을 차지해 왔다. 소수의 생물지리학자들이 과거 식물의 분포를 추적하거나 화분과 같은 프록시 자료를 통해 고기후를 복원하는 노력을 기울인 사례가 두드러진다(공우석, 2003; Park et al., 2019).
지구 시스템을 구성하는 4대 권역 중 생물권에 대한 관심의 부재는 우리나라 자연지리학의 깊이와 범위에 단순히 산술적으로 1/4만큼의 공백만 존재함을 의미하지 않는다. 자연지리학의 본질은 각 권역이 서로 고립되지 않고 물질과 에너지의 교환 및 되먹임(feedback) 작용을 통해 활발히 상호작용함을 인식하는 데 있기 때문이다(김대현, 2021; Phillips, 2021; Strahler, 1980). 이러한 상호작용과 되먹임을 통해 나타나는 자연의 창발성(emergent property)은 전체가 부분의 합보다 큼을 시사하며, 각 권역에 대한 분절적인 탐구만으로는 이해할 수 없는 우리 학문의 핵심이다(Haggett, 2001; Levin, 1998). 지구 온난화로 인해 극지방의 빙하가 녹고 해수면이 상승하면, 과연 인류는 해안습지를 잃게 되는 것일까? 이는 기권과 수권, 암석권에 국한된 사고방식일 뿐이다. 해안에 서식하는 여러 식물종은 침수가 되면 조류의 속도를 낮추고 퇴적률을 높이는 대표적인 생태계 엔지니어(ecosystem engineer)이다(Bouma et al., 2005; D’Alpaos et al., 2012; Fagherazzi et al., 2012; Kirwan and Mudd, 2012; Marani et al., 2013). 이러한 점을 명시적으로 고려하는 최신 연구들은 식물이 해안습지의 성장과 유지를 촉진함으로써 기후변화와 해수면 상승에 능동적으로 대처함을 보여준다(Kim and Lee, 2022; Kirwan et al., 2016).
본 논문은 우리나라 지리학계에 생물지형학을 정식으로 소개한 김대현(2021)의 내용을 확장한다. 이 선행 논문은 (i) 생물지형학의 정의, 역사, 주요 개념을 소개하였고, (ii) 생물지형학의 주요 용어를 한국어로 번역하는 방안을 논의하였으며, (iii) 우리나라 지리학 분야 주요 학술지에서 생물지형학적 연구의 동향을 파악하였고, (iv) 우리나라의 기후・지질 조건에서 진행할 수 있는 생물지형학적 연구의 가능성을 탐색하였다. 궁극적으로는, 한국의 지리학자들이 앞으로 이 학문 분야에 큰 관심을 기울이는 계기를 마련하고자 하였다. 이런 과정을 통해, 김대현(2021)은 생물지형학적 접근과 개념이 자연지리학의 전통에 큰 기여를 할 수 있음을 언급하였고, 생물지형학은 지형학, 생태학, 진화생물학을 아우르는 총체적 과학이라 결론지었다.
이 결론에 이어, 본 논문은 생물지형학의 학문적 근원을 이해하고 향후 나아갈 바를 모색하기 위해 다음과 같은 세 가지 세부 목적을 다룬다. 첫째, 자연지리학이 가지는 ‘복합과학’(composite science)1)으로서의 성격을 논의한다. 토마스 쿤(Kuhn)의 ‘범례’(範例, exemplar) 개념을 바탕으로, 자연지리학이 기초과학에서 차용한 원리를 통해 발전해 온 역사를 조망한다. 둘째, 자연지리학의 세부 혹은 연관 분야에서 생물지형학의 태동에 영향을 준 인물들의 학문사적 발자취를 파악한다. 각 분야의 고전으로 남은 주요 성과물들이 현재와 미래의 생물지형학에 제공하는 함의를 확인한다. 셋째, 자연지리학자들이 진화생물학적 시각을 채택함으로써 진행할 수 있는 생물지형학적 연구의 가능성을 탐색한다. 김대현(2021)에서는 깊게 다루지 못한 진화생물학적 관점, 특히 생물과 환경의 공진화 되먹임 작용을 연구하기 위한 가설을 제안한다. 궁극적으로, 우리나라 자연지리학의 가치와 지평이 생물지형학적인 연구를 포용・확장함으로써 고양될 수 있음을 강조하고자 한다.
2. 복합과학으로서의 자연지리학
1) 기초학문분야 범례의 발전과 차용
자연지리학자들은 지구 시스템을 구성하는 기권, 수권, 암석권, 생물권에 대한 포괄적인 지식을 추구한다. 그러므로 자연지리학은 기상・기후학, 수문학, 지형・지질학, 토양학, 생물・생태학 등의 다양한 학문적 원리를 바탕으로 한다. 그러나 이 사실만으로는 자연지리학이 가지는 복합학문으로서의 성격을 정확히 이해하기 어렵다. 복합학문을 규정하기 위해서는 여러 학문의 이론을 단순히 결합하는 것 이상의 의미가 있어야 한다.
『과학혁명의 구조』 제1판(Kuhn, 1962)에서 ‘패러다임’ 개념이 매우 애매하게 쓰였다는 비판에 직면하였던 토마스 쿤은 제2판에서 대체 용어로 ‘범례’를 제안하였다(Kuhn, 1970). 여기서 범례는 정상과학(normal science) 시기에 새로운 난제가 생길 시, 과학자 집단이 함께 공유하게 되는 성공적인 문제 해결 사례를 뜻한다2). 쿤은 과학사에서 패러다임의 전환이 범례의 확장, 즉 난제를 해결하는 가능성이 반복적으로 입증되는 것에 기인한다고 주장하였다. 과학자 집단의 예비 구성원들은 공유된 예제(shared example)를 구체적으로 푸는 과정을 통해 자신들의 연구가 어떻게 수행되어야 하는지 체득한다. 과학에서 중요한 발견은 문제 해결 방식을 반복하고 서로 본뜸으로써 이루어지는 것이다. 쿤의 패러다임 (혹은 범례) 개념은 정상과학의 연구 전통이 지속됨을 의미한다. 패러다임은 기존의 예측과 밝혀지는 사실 사이의 괴리가 줄어들면서 지속되지만, 시간이 지나면서 그 유용성에 반하는 사실들이 새로이 밝혀지면서 불안정해진 패러다임은 결국 새로운 것에 의해 대체된다. 새로운 패러다임으로의 대체는 기존의 학문적 전통에 대한 오랜 도전의 결과이다.
Osterkamp and Hupp(1996)은 범례를 바탕으로 기초과학과 복합과학을 구분하고 이 둘이 매우 다른 발전 과정을 보인다고 주장하였다. 기초과학은 범례를 정의하고 교체하는 과정을 통해 발전하지만, 복합과학은 적용 가능한 범례를 생성하기에는 그 자체가 너무 복잡하여 기초과학에서 차용한 원리를 바탕으로 (때로는 이들을 조합하여) 발전한다. 쿤은 물리학과 화학, 생물학을 대표적인 기초과학으로 간주하였다. 이 분야들은 만유인력의 법칙(Newton, 1687), 열역학 법칙, 화학 반응의 평형 이론(van’t Hoff, 1884), 아보가드로 법칙(Avogadro, 1810), 유전 법칙(Mendel, 1865), 자연선택 이론(Darwin, 1859)과 같은 나름대로의 범례를 개발・발전시켜 정상과학으로서의 흥망성쇠를 거듭해 왔다. 반면, 자연지리학은 기상・기후학, 수문학, 지형・지질학, 토양학, 생물・생태학을 아우르는 전형적인 복합과학으로서 기초과학의 범례를 다수 차용하여 지구 시스템의 작동 원리를 공간적인 관점에서 재해석한다. 사실, 기권, 수권, 암석권, 생물권에서 발생하는 프로세스와 상호작용을 관통하는 핵심 범례, 즉 모범적인 상부 구조가 없다 보니, 기초과학의 시각에서는 자연지리학의 방향성이 모호한 것으로 여겨질 수도 있다. 아울러, 어떤 범례는 지구 시스템의 특정 권역을 이해하는 데 유용할 수 있으나 다른 권역에 잘못 도입되어 해당 세부 분야의 발전을 저해할 수도 있다. 19세기 후반과 20세기 초반 사회사상과 과학철학에 걸쳐 전방위적으로 만연하였던 찰스 다윈(Darwin)의 진화론적 사고를 예로 들 수 있다. 이 사조의 영향을 강하게 받은 대표적인 지형학자 데이비스(Davis)와 생태학자 클레멘츠(Clements)는 자연 경관의 공간적 다양성을 인정하지 않고 각각 준평원(peneplain)과 극상(climax)으로 수렴하는 발달 원리를 기술함으로써, 시간에 의존하여 관심 현상을 설명하려는 경직된 학풍을 주도하였다(Strahler, 1952).
2) 비선형 동역학과 시스템 이론
(1) 개념 소개
본 절에서는 기초과학의 유명한 범례인 비선형 동역학(nonlinear dynamics)과 시스템 이론(system theory)을 소개하고 이 두 개념이 자연지리학적인 연구에 어떻게 적용되고 결합될 수 있는지 논의한다. 자연지리학자들은 오랜 시간 경관의 진화(landscape evolution) 과정을 밝히는 데 큰 노력을 기울여 왔다(Phillips, 1999a, 2004, 2021). 이를 통해, 지표 시스템의 (불)안정성, (비)평형, 혼돈(chaos), (비)예측성 등의 개념들이 우리 학문사를 관통하는 주제가 되었다. 이들을 야기하고 포괄하는 것이 경관의 비선형 동역학이다. 예를 들어, ‘불안정한 혼돈의 시스템(unstable chaotic system)’ 이론은 발산적 경관(divergent landscape) 개념에 적용되어 미세한 기후・지질 조건의 차이가 시간이 지남에 따라 증폭되어 지역별로 지형 경관의 큰 변이를 불러옴을 설명한다(Kim et al., 2016a). 한편, 비선형성은 안정적인 비혼돈 시스템(stable nonchaotic system)에도 적용되어 초기 조건의 지역별 변이가 감소하여 수렴적 경관(convergent landscape)이 발현하는 경우도 포함한다. 이러한 비선형성을 고려하지 않는다면, 지역별로 존재하는 다양한 생물적・비생물적 차이가 증가하거나 감소하지 않고 오랜 시간 그대로 지속된다는 비현실적인 예측이 야기될 것이다. 비선형 동역학은 지표 시스템 전반에 걸쳐 흔하게 작동하고 있으며, 자연지리학의 전통에 통합적 틀로서 기능할 여지가 충분하다(Kim, 2010, 2017; Phillips, 2003; Thornes, 2003).
‘시스템’은 오스트리아의 생물학자 폰 버탈란피(von Bertalanffy)가 1950년대에 일반 시스템 이론을 제창한 이후(von Bertalanffy, 1950, 1951), 아마 자연지리학계에서 가장 흔하게 쓰여 온 단어일 것이다. 특히, 1980년대부터 ‘비선형 동역학 시스템(nonlinear dynamic system, NDS)’ 개념이 경관 진화 연구에 활발히 사용되기 시작하였다(Culling, 1987; Kirkby, 1995; Phillips, 1999b; Thornes, 1985). 다만, 자연지리학 분야에서 시스템 이론에 비선형 동역학 개념이 통합된 배경에 대해서는 진지한 고민이 이루어지지 않은 것도 사실이다. 과학자들이 둘 사이에 진정 긴밀한 연관성이 존재함을 밝혀낸 것인지, 혹은 시스템 앞에 마치 유행처럼 새 개념을 가져다 붙여 특수용어를 만들어 내는 일환(Sherman, 1996; Thomas and Goudie, 2000)에서 NDS가 생겨났는지 명확하지 않다.
(2) 자연지리학에서의 시스템 이론
자연지리학자들은 전통적으로 환원주의적 접근과 총체주의적 접근이 서로 상충하는 데 관심을 가져왔다(Bauer et al., 1999; Malanson, 1999; Phillips, 1999b; Strahler, 1992). 환원주의적 관점은 자연 경관을 몇 개의 부분으로 나누어 자세하게 분석(analysis)한다. 이 부분들의 합은 해당 경관이 된다. 총체주의적 접근은 각 부분에 대한 세부적인 분석보다는 종합(synthesis)에 주안점을 두고 전체 경관의 작동 원리에 대한 큰 그림을 그리고자 한다. 많은 자연지리학자들이 채택하는 시스템 접근에서는 분석과 종합 모두 중요한 요소이다. 시스템은 서로 상호작용하는 여러 부분들의 조합으로 정의된다. 모든 시스템은 아래와 같은 몇 가지 공통점을 지닌다(White et al., 1992).
(i) 일련의 구조 혹은 조직을 갖춘다.
(ii) 실제 자연을 어느 정도 일반화 혹은 추상화한다.
(iii) 일정한 기능을 보인다.
(iv) 그러므로 구성 요소들 사이에 구조적이고 기능적인 관계가 존재한다.
(v) 기능은 물질과 에너지의 이동 또는 교환을 의미한다.
(vi) 이 기능을 작동하는 주요 힘 혹은 에너지의 원천이 존재한다.
(vii) 구성 요소들이 통합되어 하나의 시스템을 정의한다.
시스템은 외부와의 에너지와 물질의 교환 여부에 따라 크게 고립된 시스템, 닫힌 시스템, 열린 시스템으로 나뉜다. 고립된 시스템은 시스템과 외부 환경 사이에 아무런 상호작용이 존재하지 않는 경우이다. 닫힌 시스템의 경우, 외부와 물질 교환은 없지만 에너지가 이동할 수는 있다. 실제 세계에서 고립되거나 닫힌 시스템의 예를 찾기는 어렵다. 열린 시스템은 외부와 물질과 에너지를 원활하게 교환한다. 모든 자연 시스템은 이 경우에 해당하며 열역학 법칙을 따른다(White et al., 1992). 자연지리학의 선구자들은 대자연을 역동적인 열린 시스템으로 간주하고, 기권, 수권, 암석권, 생물권으로 나누어 이들 사이의 상호작용, 즉 물질과 에너지의 교환을 살펴봄으로써 지역적 유사점과 차이점을 규명해왔다(Chorley, 1962; Krumbein, 1963; Odum, 1972; Strahler, 1952, 1980).
시스템 이론은 생물학에 본격적으로 도입되기 이전에는 물리학과 화학 분야에서 닫힌 반응 시스템(closed reaction system)의 프로세스를 설명하는 데 주로 사용되었다. 그러나 생명체는 물질과 에너지가 원활하게 흐르는 열린 시스템이기 때문에, 전통적인 닫힌 시스템 사고와 생물학의 열린 시스템 사고 사이의 상반된 특성을 고려할 필요가 있었다. 이를 위해 폰 버탈란피는 등결과성(equifinality)3) 과 열역학(thermodynamics)을 거론하였다. 등결과성은 시스템의 최종 상태나 모습이 상이한 초기 조건과 과정을 통해서도 발현될 수 있음을 뜻한다. 이를 통해, 닫힌 시스템과 열린 시스템 사이의 근본적인 차이를 설명할 수 있다. 닫힌 시스템에서는 최초 조건이 최종 상태를 결정하기 마련이다. 예를 들어, 화학적 평형 상태에서는 반응물의 최종 농도는 최초 농도의 수준에 따라 결정된다. 만약 최초 농도나 화학 반응 프로세스에 변화가 생긴다면 최종 농도는 이에 따라 변할 것이다. 그러므로 닫힌 시스템에서는 등결과성을 기대할 수 없다. 열린 시스템은 그렇지 않다. 간단한 예로, 다 자란 성게의 모습은 하나의 완전한 난자에서 비롯되었건, 반으로 나뉜 난자 각각에서 비롯되었건, 합쳐진 두 개의 완전한 난자에서 비롯되었건 동일하다. 열린 시스템에서는 외부와의 물질 교환이 자유롭기 때문에 초기 조건의 변이와 무관하게 동일한 결과를 예상할 수 있는 것이다. 자연지리학자들은 상이한 초기 조건과 지표 프로세스를 통해서도 동일한 지형 경관이 야기되는 많은 사례를 발견해왔다(Beven, 1996; Chorley, 1962; Culling, 1957, 1987; Haines- Young and Petch, 1983). 연흔(ripple)은 실트로 구성된 조간대에서 조류의 작용으로 혹은 해안사구에서 바람에 의해 형성될 수 있다.
닫힌 시스템에서의 열역학 법칙을 열린 시스템에 적용하는 경우에도 상당히 다른 결과가 도출된다(von Bertalanffy, 1950). 특히, 열역학 제2법칙은 닫힌 시스템의 시간-의존적(time-dependent)인 평형 상태에서만 적용되는 것으로 알려져 있다. 에너지는 결국 높은 곳에서 낮은 곳으로 흐르며 자연의 프로세스는 비가역적이고 과거와 미래의 상태는 서로 비대칭적일 수밖에 없다. 그 어떤 에너지 흐름도 완벽하게 효율적일 수는 없고 일부는 소산되기 마련이다. 엔트로피는 이 소실되는 에너지의 양을 의미하며 시스템 무질서의 표현이기도 하다. 자연지리학에서는 경관 진화에 대한 데이비스의 이론(Davis, 1899)이 닫힌 시스템의 열역학 개념과 맞닿아 있다(Chorley, 1962). 지반의 융기는 지표 시스템에 일정 수준의 위치에너지를 부여한다. 삭박(denudation)이 진행되면 이 에너지가 점진적으로 감소한다. 최종적인 준평원 단계에서는 기복을 잃은 지표면이 최소한의 자유 에너지를 지니는 최대 엔트로피 상태에 접어든다. 이것이 닫힌 시스템에서의 비가역적인 경관 진화 과정이다. 사실 열역학 제2법칙을 열린 지표 시스템에 이렇게 직접 적용하는 것은 바람직하지 않다. 물론 열린 시스템도 꾸준한 비가역적 프로세스로 인해 증가하는 엔트로피를 피할 수는 없다. 그러나 열린 시스템은 외부 환경과의 물질과 에너지 교환을 통해 최대 엔트로피, 즉 열역학 평형 상태에 이르지 않고 동적 정상 상태(dynamic steady-state)를 유지한다. 예를 들어, 사면 시스템은 삭박을 통해 경사가 낮아지더라도 상부 사면으로부터 새로운 물질을 공급받아 (이를 통해 자유 에너지가 보충됨) 기존의 안식각을 유지할 수 있다.
(3) 자연지리학에서의 비선형 동역학
물리학은 갈릴레오 시대 이후 20세기 초반까지 눈부신 성장을 이루었다. 이 배경에는 에너지와 물질의 투입(input)과 결과(output) 사이에 존재하는 비례 관계, 즉 선형성(linearity)에 바탕을 둔 자연 현상의 해석이 있다. 선형성이 만족된다면, 전체 시스템을 여러 부분으로 나누어 연구하는 단순화된 환원주의적 접근에 큰 문제가 없을 것이다. 자연을 점점 더 작은 단위로 쪼개어 살펴보고자 하는 과학자들의 시도가 수백 년간 이어졌다. 세포에서 분자로, 분자에서 원자로. 이러한 선형성 패러다임은 20세기 후반 비선형의 과학으로 대체되었다(Strogatz, 2003). 특히 부분의 합이 전체보다 클 수 있다는 논의는 창발(emergence) 혹은 창발성 개념의 대두로 이어졌다. 창발은 구성 요소(하위 계층) 단위에서 존재하지 않지만, 요소들 사이의 상호작용의 결과 전체 시스템(상위 계층)에서 돌연히 출현하는 특성 또는 현상을 의미한다(Johnson, 2001). 이렇게 나타나는 시스템의 비선형성은 물질과 에너지의 입력과 출력 사이에 비례 관계가 성립하지 않음을 의미한다.
선형성과 비선형성의 논의를 수학적으로 이어가 보자. 어떤 함수가 선형성을 띤다는 것은 변수들의 가중치 합이 함수값과 동일함을 의미한다: f(ax+by) = af(x)+bf(y). 기하학적인 관점에서는 선형 함수를 일직선으로 나타낼 수 있다: y = mx + b (m: 기울기, b: y절편). 한편, 비선형 함수는 x가 다른 변수나 자기 자신과 곱해지는 경우로 표현할 수 있다. 후자의 경우를 멱함수(power function)라고 한다. 생물학의 전통적인 포식자-피식자 관계가 비선형 함수의 예가 된다. 포식자와 피식자 중 어느 하나의 개체군(population) 크기가 커진다면 두 집단이 마주칠 확률이 높아질 것이다: Z = cUV (Z 단위 시간당 마주치는 횟수, U 포식자 마릿수, V 피식자 마릿수, c 포식자의 사냥 효율성을 나타내는 상수). 이 함수는 두 개의 변수가 더해지지 않고 곱해짐으로써 정의되기 때문에 비선형 관계를 가진다(Holland, 1995).
비선형성은 지표 시스템에서 흔하게 발견된다(Phillips, 2003; Thornes, 2003). 일찍이 Brunsden(1990)과 Lane and Richards(1997)는 비선형 지형학이 자연지리학과 경관 진화 모델링의 미래가 될 것으로 내다보았다. 혼돈 이론에서는 특정 기후대 안에서도 지역별로 나타나는 미세한 지질・수문・토양・식생의 차이가 시간이 지나면서 증폭되어 발산적 경관이 야기된다고 예측한다. 그러나 동시에 혼돈의 동역학 개념에 의하면, 확연한 무질서 속에 내재된 질서, 즉 결정론적 혼돈(deterministic chaos)이 존재하기 때문에 경관의 발산이 진행되더라도 이는 결정론적이며 유한하다(Phillips, 1999a).
3) 비선형 동역학과 시스템 이론의 연결
열린 시스템은 물질과 에너지의 흐름을 통해 정상 상태에 접어듦으로써 등결과성을 유지할 수 있다(von Bertalanffy, 1950). 이런 흐름이 없다면, 열린 시스템 안에서 발생하는 현상(예: 체온)의 평균값 근처에서는 역동적인 변화가 나타나기 어렵다. 다시 말해, 열린 시스템은 거시적 스케일에서는 완전한 정적 평형(static equilibrium)처럼 보일 수 있겠지만 미시적 스케일에서 자세히 관찰하면 그렇지 않다. 열린 시스템이 이러한 동적인 정상 상태를 보이는 것은 평균 상태에서 크게 벗어나지 못하게 억누르는 음의 되먹임(negative feedback)을 통한 자기 조절(self-regulation) 능력 때문이다. 우리가 운동을 통해 체온이 상승하더라도 땀을 흘림으로써 항상성을 유지하는 것과 같은 이치다. 열린 시스템은 바로 이 자기 조절을 통해 초기 조건이나 외부 교란(perturbation), 기타 프로세스 등에 상관없이 등결과성을 보인다. 여기서 중요한 점은 열린 시스템의 어떤 상태도 반드시 평균값에 일치하지 않고 그 위 아래 가까이에서 변동할 뿐이라는 것이다(Culling, 1987; White et al., 1992). 이는 이력(hysteresis) 개념과 연관된다. 이력은 시스템의 속성이나 현상이 이전 상태와 완벽하게 동일한 모습을 띨 수 없음을 의미한다. 예를 들어, 어떤 하천생태계에서 홍수 이전과 이후에 기온, 강수, 토양, 지형 등의 환경 조건이 동일하게 주어졌다고 가정하더라도, 홍수 이후 하천생태계가 재생하는 과정은 교란 이전에 그 생태계에서 진행되었던 천이(succession)의 과정과 동일할 수 없다. 홍수 직후 정착하는 식물종의 조성이 홍수 이전의 그것과 분명히 다를 것이기 때문이다.
(1) 이력의 비선형적 속성
이력은 단일 투입값 혹은 설명 변수값에 대해 둘 이상의 결과값 혹은 종속 변수값이 존재한다는 측면에서 열린 시스템의 비선형성을 보여준다(Phillips, 2003). 예를 들어, 수문곡선(hydrograph)은 비가 온 이후 하도 내 유량의 변화를 시간적으로 나타내는데, 첨두유량의 좌우로 각각 상승곡선과 하강곡선을 보인다. 두 곡선에서 동일한 유량을 가지는 두 시간대는 퇴적물의 양이 서로 다르다. 첨두유량으로 향하는 상승곡선 구간에서는 하상의 침식으로 인해 하강곡선 구간에 비해 더 많은 퇴적물이 운반되기 때문이다. 아울러, 다음 홍수가 발생하여 이전 홍수 때와 동일한 유량에 다다르더라도 그 순간 운반되는 퇴적물의 양은 이전 홍수의 해당 유량에 의해 운반되던 퇴적물의 양과 같을 수 없다. 이전 홍수가 발생한 이후, 하도의 지형과 식생의 분포는 크게 바뀌었을 것이기 때문이다. 이렇듯, 열린 시스템의 속성이 동일한 조건에서도 똑같이 반복되지 않고 항상 변화하는 현상은 그 시스템의 임계 변동(threshold behavior) 및 창발성에 기인한다.
시스템의 임계 변동은 지표 시스템에서 흔하게 발견되며 비선형성의 주요 원인이다(Kim, 2017; Phillips, 2003). 어떤 현상 혹은 변수가 임계치에 다다르면 시스템의 급진적인 변화가 야기될 수 있다. 지형학에서 대표적인 임계 변동의 사례는 사면의 안식각에서 찾을 수 있다. 폭우나 지진으로 인해 사면의 안정성이 감소한다면 약간의 교란에도 급격한 대규모 매스 무브먼트가 발생할 수 있다. 이는 작은 투입값이 큰 변화(결과값)를 불러온다는 점에서 시스템 비선형성의 예가 된다. 선형성에 기반한 사고방식에서는 기대할 수 없는 결과이다.
창발성 역시 외부 교란 이후 시스템의 회복 과정에 뚜렷한 비선형성을 유발함으로써(Thornes, 2003), 환원주의적 접근에 의해 생성된 분절적인 정보의 조합을 불완전하게 만든다. 음악감상의 예를 들어 보자. 우리는 관현악단의 대규모 연주를 들음으로써, 여러 악기가 내는 소리의 조화와 상호작용에 감동을 느낀다. 이는 바이올린, 첼로, 플루트, 트럼펫 등의 각 연주를 순차적으로 따로 듣고 누릴 수 있는 느낌과 다른 것이다. 관현악 합주가 주는 전체적인 감동이 단일 악기가 주는 감동의 단순한 합계가 아닌 것처럼, 기권, 수권, 암석권, 생물권 각각의 프로세스를 따로 연구함으로써 지구 시스템 전체에 대한 이해를 도모하는 것은 바람직하지 않다. 이처럼 창발성은 부분들의 합을 전체보다 크게 만든다는 점에서 시스템 비선형성의 원인이 된다(Holland, 1995; Johnson, 2001). 앞서 언급한 포식자와 피식자의 상호작용에서도 창발성을 발견할 수 있다. 이 상호작용을 이해하기 위해 U와 V 사이에 덧셈이 아니라 곱셈을 취해야 함은 이미 언급하였다. 추가적으로, 특정 시간대 t+1에 존재하는 포식자의 마릿수 Ut+1을 추정하기 위해서는 사냥한 피식자를 섭취함으로써 자손으로 전환하는 효율을 의미하는 또 다른 상수 ‘r’을 고려해야 한다: Ut+1 = Ut - dUt + bUt + r[cUtVt] (dUt: 시간대 t와 t+1 사이에 죽은 포식자의 마릿수, bUt: 시간대 t와 t+1 사이에 태어난 포식자의 마릿수). 위 식의 마지막 부분은 포식자가 피식자를 잡아먹을 때마다 생기는 창발성, 즉 더 튼튼하고 왕성한 번식력을 가지는 포식자 개체를 나타낸다.
(2) 엔트로피의 비선형적 성격
지형 경관은 서로 긴밀히 연결되어 상호작용하는 다양한 부분으로 구성되어 있다. 예를 들어, 하나의 유역(catchment)은 사면과 토양, 식물, 유수, 미기후 등으로 이루어진다. 이 유역 경관의 진화 과정은 많은 경우 외부 압력으로부터 독립적, 즉 자기 조직적(self-organizing)이다(Jørgensen et al., 1998; Phillips, 1995, 1999a). 유역의 여러 구성 요소들이 서로 상호작용하며 자발적으로 새로운 유역 경관을 창출한다는 뜻이다. 자연에서 경관의 자기 조직화는 크게 수렴형과 발산형으로 나뉜다(Kim et al., 2016a). 전자는 초기 조건의 차이와 상관없이 여러 지역의 모습이 일련의 질서와 규칙성을 지닌 동일한 형태로 변해가는 현상을 의미한다. 후자는 여러 지역의 지형 경관이 가지는 최초의 미세한 차이가 시간이 지남에 따라 증폭되어 서로 다른 모습으로 바뀌어 가는 것을 말한다.
이 두 유형의 경관 진화는 각각 최소 엔트로피와 최대 엔트로피 개념과 연관되기 때문에 자연지리학적 시스템 접근에서 중요하다(Phillips, 1999a). 수렴적 경관 진화 모델은 지표 시스템의 안정성과 엔트로피의 감소를 가정한다. 반면, 발산형 모델은 시스템의 불안정성 및 결정론적 혼돈과 연관되어 엔트로피의 증가를 예측한다. 후자의 경우가 초기 조건의 기하급수적인 증가를 의미하므로 시스템 비선형성의 좋은 예가 된다. 콜모고로프(Kolmogorov) 엔트로피(K-엔트로피), 혼돈, 불안정성 사이에는 직접적인 관계가 있다. 특히, 유한한(finite) 양(positive)의 K-엔트로피는 불안정한 혼돈을 통해 경관의 발산을 야기하므로(Phillips, 1999b) 시스템의 비선형성을 의미한다.
발산형 시스템의 자기 조직적인 비선형 동역학을 이론적으로 다시 살펴보면, 외부 교란 이후 시간에 따라 시스템이 수렴적으로 혹은 발산적으로 진화할 지 여부를 다음과 같은 식으로 예측할 수 있다: δ(t) = Ce-λt (δ(t): 시점 t에서의 외부 교란, C: 상수 혹은 초기 조건, λ: 리아푸노프 지수 혹은 고유값). 만약 λ < 0이라면, 외부 교란의 영향은 기하급수적으로 감소하여 이전 상태로 돌아갔음을 의미한다. 이는 시스템의 안정성과 수렴을 뜻한다. 그 반대의 경우(λ > 0), 초기의 변이가 시간이 지남에 따라 기하급수적으로 증폭되어 불안정성과 발산이 발생하였음을 뜻한다. 각 리아푸노프 지수의 크기는 K-엔트로피와 직접적으로 연관된다. 후자는 리아푸노프 지수들 중 양의 값들의 합과 같기 때문이다. 그러므로 리아푸노프 지수, K-엔트로피, 시스템의 비선형 동역학 사이에는 논리적인 관계가 존재함을 결론지을 수 있다. 예를 들어, 초기의 토양과 수문 조건의 차이가 시간의 경과에 따라 기하급수적으로 증가하면 식생 피복의 유형은 여러 공간 단위별로 분화된다. 이러한 경관 발산에 의해 일부 혹은 모든 리아푸노프 지수와 K-엔트로피는 양의 값을 가지게 된다.
4) 소결
전술하였듯, 비선형성이 지표 시스템에 만연해 있다는 점은 더 이상 놀라운 사실이 아니다(Kim, 2017; Phillips, 2003; Thornes, 2003). 본 절에서는 비선형 동역학 시스템(NDS) 개념이 자연지리학에 대두된 배경을 논하였다. 보다 구체적으로는, 비선형 동역학과 시스템이 어떤 이유와 논리를 통해 통합되었는지 설명하였다. 아울러, 등결과성은 이력, 임계, 창발성 등의 개념과 연결됨으로써, 비선형 동역학과 시스템 이론을 잇는 징검다리 역할을 함을 밝혔다. 엔트로피가 초기 변이의 기하급수적 증폭을 의미하는 혼돈의 측정치로서 시스템의 비선형성을 지시함도 밝혔다.
본 절에서 다룬 다양한 개념은 물리학, 화학, 생물학 등의 기초과학에서 개발・발전시켜 온 범례들이다(Kuhn, 1970). 자연지리학은 이들을 차용・혼합하여 기권, 수권, 암석권, 생물권 사이에 발생하는 복잡한 상호작용을 설명하고자 하는 복합과학이다. 자연지리학의 이러한 특성은 후술하듯이 지형학, 생태학, 진화생물학을 통합하는 생물지형학의 복합적 성격(김대현, 2021)과 정확히 맞닿아 있다. 그러므로 많은 자연지리학자들이 스스로의 관심과 전문성을 생물지형학으로 돌리는 것은 상당히 자연스럽고 독려할 만한 현상이다. 특히, 지형학을 스스로의 학문적 뿌리로 생각하는 자연지리학자가 생태학과 진화생물학을 꾸준히 깊고 넓게 접하는 분위기가 바람직하다.
3. 생물지형학의 탄생과 발달
1) 지형학, 토양학, 생태학: 학문사적 흐름의 재발견
우리나라 지리학계에 생물지형학이 소개된 이후(김대현, 2021), 염습지 생태계(박건우 등, 2023)와 하천생태계(이건학 등, 2024; Choi, 2024; Lee et al., 2023)를 중심으로 생물지형학적 개념과 접근을 바탕으로 한 새로운 연구가 진행되고 있다. 본 절에서는 생물지형학의 배경이 되는 주요 분야들의 학문사적 흐름을 살펴봄으로써, 한국 지리학계 내에서 생물지형학에 대한 이해의 지평을 넓히고자 한다. 생물지형학은 자연지리학의 여러 내용을 바탕으로 하는 정수(精髓)로서, 자연지리학의 세부 분야들이 교차하는 지점에서 새로운 지식을 창출하고 있다. Corenblit et al.(2008)은 지형학, 생태학, 진화생물학이 지니는 한계를 바탕으로 생물지형학이 태동했다고 설명한다. 지형학에서는 지형과 관련된 역학적 프로세스를 생물체의 활동과 독립적인 것으로 여겼다. 생태학에서는 서식처와 생물의 관계를 연구함에 있어 생물이 서식처에 미치는 능동적인 영향을 포함하지 못하였다. 마지막으로, 진화생물학에서는 물리적 환경에 대한 생물의 영향과 그 결과적인 되먹임을 고려하지 못하였다. 따라서 생물지형학의 근원을 파악하기 위해서는 지형학, 생태학, 진화생물학의 학문사적 흐름을 살필 필요가 있다. 특히, 자연지리학의 맥락에서는 지표 시스템을 다루는 핵심 분야인 지형학, 토양학, 생태학의 발달 과정을 깊게 조망해야 한다. 이 분야들은 19세기 후반과 20세기 초반을 기점으로 독립된 학문으로 발전하기 시작하였다. 각 학문 분야의 초기 이론가들은 주로 문헌을 통한 간접적인 방식으로 상호 간 영향을 미쳤다. 그러나 20세기 후반까지 이들을 통합한 실질적인 학제 간 연구는 거의 이루어지지 않았다. 특히, 패러다임(범례), 관점, 연구 방법 등의 차이로 인해 명백한 유사성을 가진 세 학문의 융합이 어려울 수밖에 없었다(Hupp et al., 1995; Renschler et al., 2007).
한편, Osterkamp and Hupp(1996)에 따르면 지형학과 토양학, 생태학에서 공통적인 연구 흐름이 특정한 순서로 발전했다. 이 흐름은 (i) 관찰과 기술을 중점적으로 다루는 관찰론자(observationalist), (ii) 찰스 다윈의 『종의 기원』으로부터 진화론을 차용한 다윈주의자(Darwinist), (iii) 화학의 평형 패러다임을 채택한 평형론자(equilibrist), 그리고 (iv) 진화 패러다임과 평형 패러다임을 통합하여 새로운 관점을 부여한 통합론자(integrationist)로 구분된다(표 1). 관찰론자와 다윈주의자, 평형론자, 통합론자 모두 개별 학문에서의 패러다임과 관점의 전환을 다루는 경향을 보인다. 이 흐름의 끝으로는 융합론자(unifier)가 제시되었는데, 본 논문에서는 통합적인 관점을 기반으로 학제 간 융합을 구현한다는 점에서 생물지형학자를 융합론자로 본다.
표 1.
Osterkamp and Hupp(1996)를 기반으로 재구성한 연구 사조 정리
2) 각 학문사적 발전 과정이 생물지형학의 태동에 미친 영향
지형학, 토양학, 생태학에서 다루는 대상은 서로 근본적으로 다르다. 그럼에도 불구하고 이들 사이에는 약속이나 한 듯 비슷한 시기에 나타나는 공통된 흐름이 존재한다. 본 절에서는 이 흐름을 정성적 기술 및 연역적 접근, 개념적 차원의 동역학적 접근, 정량적 객관화와 실증주의적 접근, 시스템 이론과 복잡계적 접근 등 네 가지로 요약한다. 이들은 대체로 시간적 연속선상에 배치된다.
이 네 가지 흐름을 간략히 소개하면, 학문 초기 단계에는 정성적 기술과 연역적 접근을 통해 연구가 진행되었다. 특히 현장 답사는 자연환경을 직관적으로 이해하는 효과적인 방법이었다. 연구자들은 답사를 통해 자연 현상을 직접 관찰하고 그 특성과 과정에 대해 상세히 기술하였다. 한편, 당시에는 일반적인 원리에서 개별적인 사례를 이해하려는 연역적 접근을 바탕으로 한 연구가 주를 이뤘다. 진화론의 영향을 받아 시간의 흐름에 바탕을 두고 자연 현상을 확인하려는 경향도 나타났다. 앞서 언급한 데이비스와 클레멘츠의 경우처럼, 특정 이론이나 원리에 갇힌 상태로 장기간에 걸쳐 경직된 학문적 논의가 진행된 경우에는 해당 학문의 발전에 큰 저해 요소가 되기도 하였다. 결국, 한 가지 원리를 통해 모든 현상을 설명하는 것에는 한계가 있었으며, 관심 현상과 관련된 다양한 요인들의 상대적 영향력을 고려하는 동역학적 접근이 나타났다. 자연 현상에는 다양한 요인이 작용하고 있으며 각각의 요인은 서로 다른 영향력을 가질 수 있다는 사실을 인정하였고, 개념적 차원에서의 논의를 통해 관심 현상을 이해하기 위한 틀이 마련되었다. 이러한 과정에서 타학문과의 연결을 통해 해답을 찾기도 하였다. 20세기 중반 계량 혁명을 거치면서, 관심 현상을 수량화된 자료와 통계적 방법을 통해 파악하는 정량적 객관화와 실증주의적 접근으로 이어졌다. 이를 통해, 연구 결과를 수치화하고 통계적 유의성을 평가할 수 있었으며, 연구의 정확성과 객관성을 확보할 수 있게 되었다. 한편, 자연 현상을 바라보았던 과거의 시선은 조화와 균형, 질서와 안정을 이루는 아름다운 집합체였다. 그러나 실제 자연 시스템은 구성 요소의 단순한 합으로 표현하기 어려우며, 이들 사이의 복잡한 상호작용을 통해 비선형적인 반응을 보인다. 아울러, 시스템은 교란으로 인해 안정적인 평형 상태로 도달할 수 없다는 비평형(non-equilibrium)적 관점이 부각되었다. 이러한 흐름 속에서 구성 요소 간 상호 의존성과 비선형성을 전제로 하는 시스템 이론과 복잡계적 접근이 대두되었다.
아래 이어지는 네 개의 소절에서는 각 흐름을 보다 자세히 정리한다. 특히, 지형학, 토양학, 생태학의 주요 인물들과 이들의 고전 논문을 소개하며, 각 분야의 학문사적 발달 과정이 생물지형학의 태동에 미친 영향과 통찰을 (i) 학제 간 연결과 융합, (ii) 다중 시공간 스케일 관점과 범주의 다양화, (iii) 비선형성을 토대로 하는 복잡계적 시각의 적용 등 세 가지 측면에서 논의한다. 성공적인 학제 간 융합은 ‘두 개 이상의 영역을 하나의 개념적・경험적 구조로 뚜렷이 결합하는 것’(Pickett et al., 1994; Renschler et al., 2007)이다. Renschler et al.(2007)는 지형학과 생태학을 결합한 연구에서 자주 발생하는 문제가 스케일임을 명시한 바 있다. 특히, 생물-지형 복합체(organism-landform complex)에 적용되는 다양한 되먹임 양상은 둘 이상의 다중 범주를 기반으로 한다(Thoms et al., 2018; Wheaton et al., 2011). 따라서, 세부 학문 분야간 온전한 통합과 융합을 기반으로 하는 생물지형학에서는 다양한 스케일 범주에 대한 고민이 꾸준히 수행되어야 한다. 한편, 생물지형학과 관련된 기존 연구는 지형과 생물적 요소의 비선형성을 인지하고 있다(Hession et al., 2010; Renschler et al., 2007; Thoms et al., 2018; Wheaton et al., 2011). 많은 학계에서 이미 평형 패러다임을 넘어 비평형 패러다임을 제기했으며, 생물-지형 복합체의 특성 또한 비선형성과 복합성을 바탕으로 설명해야 한다.
(1) 정성적 기술 및 연역적 접근
정성적 기술과 연역적 접근의 맥락에서 지형학의 대표적인 학자는 데이비스이다. 그는 지형의 발달을 거시적으로 설명하는 틀로서 지형윤회설을 주장하였다(Davis, 1899). 다윈의 진화적 관점에 착안하여 생명체의 생애주기와 같이 지형 경관의 발달을 시간의 진행에 따른 다단계 모델로 설명하고자 하였다. 한편, 생태학에서는 식생의 발달 과정이 궁극적으로 최종 상태인 극상에 도달한다고 설명하였다(Clements, 1916; Phillips, 1934). 클레멘츠의 천이 이론에서 여러 식물 종들은 하나의 초유기체처럼 존재하며, 군집(community)으로서 특정 기후 조건을 대변한다.
데이비스와 클레멘츠는 지형과 생태 경관의 다양성을 인정하지 않고, 각각 준평원과 극상으로 수렴하는 발달 원리를 통해서 실제 자연의 광범위한 경관을 기술적으로 설명하였다. 당시의 과학철학은 다윈의 진화론을 바탕으로 형성되어 있었기 때문에, 시간 흐름에 의존하여 관심 현상을 설명하는 학풍이 만연하였다. 또한, 측정 기술이 발달하지 않았기에 연역적 추측을 바탕으로 연구가 진행되었으며, 방어적이고 현학적인 글쓰기가 이루어졌다. 결국, 단일 원리를 통해 광범위하게 나타나는 현상을 설명하는 것에는 한계가 있었다. 이들의 이론은 후대 학자들에게 많은 비판을 받았지만, 초기 자연과학의 학문적 토대를 마련하였다는 점은 분명하다.
(2) 개념적 차원의 동역학적 접근
지형학에서 관심 현상과 관련된 다양한 요인들을 고려한 동역학적 접근의 선구자는 길버트(Gilbert)이다. Gilbert (1909)는 산정부의 볼록한 사면이 형성되는 과정을 토양포행을 통해 설명했으며, 그에 영향을 미치는 요인으로 토양 입자의 팽창과 수축, 동식물, 바람, 강우 등을 고려하였다. 데이비스의 논의는 역사적인 관점에서 이루어졌기 때문에, 구조나 프로세스는 지형 발달 단계에 비해 덜 중요하게 여겨졌다. 이와 대조적으로, 길버트는 균형, 역학적 조정, 평형을 강조하여 구조와 프로세스를 설명하였다(Morisawa, 1988).
한편, 여러 토양학자들은 기후, 유기체, 지형, 모재, 시간 등 다양한 요소를 통해 토양 시스템을 설명하였다(Dokuchaev, 1948; Thorp, 1941). Milne(1935)는 모재의 조건뿐 아니라 지형적 관계를 동시에 고려하여 토양을 분류하는 카테나(catena) 개념을 제안하였다. 그의 연구에서는 카테나가 지도화를 위한 단위로 제시되었지만, 이 개념은 토양을 지형적 맥락에서 이해한 첫 시도이다. 이를 통해, 모재나 형태적인 차이가 존재하더라도 지형 조건과 토양이 일련의 관계를 보일 수 있으며, 사면에서의 위치에 따라 토양 속성에 영향을 미치는 조건들 사이의 상호작용이 다르게 나타남을 암시하였다. Bryan and Albritton(1943)은 기후의 영향으로 인한 토양의 공간적 변이를 설명하였다. 기후의 영향을 받은 생물, 화학, 물리적 작용과 더불어 시간과 인간 활동의 영향으로 토양 성질의 차이가 나타난다고 주장하였다. 또한, 토양을 통해 과거 기후변화를 해석하고자 하였으며, 지질 사건과 토양 사건 사이의 상관관계를 확인하여 토양 형성 프로세스에 다양한 요인이 작용할 수 있음을 확인하였다. 토양의 기원을 지질 연대 및 부정합과 같은 지질・지리학적 증거와 연결 지어 해석했다는 측면에서, 학제 간 연결과 융합이 구현된 고전 논문으로 여겨진다.
생태학 분야에서는 식생 분포를 설명하기 위한 노력이 개념적 차원의 동역학적 접근으로 이어졌다. Cowles(1899)는 미시간 호수 현장 답사를 바탕으로 사구 식생의 분포를 설명하였다. 식생은 과거와 현재 환경 조건의 산물이며, 식생의 분포는 물리 및 생물 요인의 영향을 크게 받는다고 논의되었다. 그 예시로 빛과 열, 바람, 토양, 물, 그리고 기타 요소로서 산불, 매연, 지형, 동물, 인간 등이 식생 분포에 미치는 영향 등이 거론되었다. 코울스(Cowles)의 1899년 논문은 시간의 흐름과 공간적 분포를 적용한 생태학의 고전 논문으로 여겨진다. 생물학자였던 그는 전반적인 경관 변화에 내재된 식물 군락의 연속적인 순서와 이를 지배하는 법칙을 지형 요소를 고려하여 설명하였다. 이를 토대로, 식물 군락이 ‘두 번 다시 동일하게 반복되지 않는 파노라마(panorama, never twice alike)’임을 역설하였다. 여기서 식생 군락은 유사한 지형과 서식지를 공유하고, 비슷한 환경 조건에 노출된 식물들의 군집으로 정의된다. 이는 상대적으로 불규칙한 조각(patch)과 비교적 대칭적인 구역(zone)으로 세분화될 수 있다. 코울스는 다윈과 데이비스의 영향을 받았으나 시간의 흐름에 따른 식생 천이를 논의하는 데 그치지 않았다(Osterkamp and Hupp, 1996). 그는 더 나아가 식생 군락이 지형과 다양한 환경 조건의 영향을 받음을 명시함으로써 시공간적 범위를 복합적으로 고려한 최초의 생태학자로 평가된다. 또한, 식생 군락의 공간적 분포를 조각과 구역으로 세분화하였다는 측면에서, 후대 생태학자들과 생물지형학자들의 공간 스케일적 사고에도 영향을 주었다. Gleason(1926)과 Ramensky(1930) 역시 환경 요인이 식물 분포에 미치는 영향을 연구하였다. 식물 군집을 유기적인 전체로 보았던 클레멘츠와 다르게, 이들은 식생의 분포를 개별 식물종이 확산・이동하고 환경 조건의 영향을 받은 결과로 해석하였다. 특히, 라멘스키는 개별종의 분포와 군집의 관계까지 논의를 확장하여, 후술하게 될 구배 분석(gradient analysis)의 창시자가 되었다(Whittaker, 1967).
본 소절에서 언급된 학자들은 단계적인 발달 과정을 통해 관찰 결과를 연역적으로 서술하던 이전의 흐름에서 벗어나, 다양한 요인과 그 영향력의 상대적 중요도까지 포함하여 관심 현상을 설명하는 개념적 틀을 제시하였다. 요인 간의 상호작용을 고려한 이들의 논의는 자연과학이 오늘날과 유사한 모습으로 진전하는 데 중요한 역할을 하였다.
(3) 정량적 객관화와 실증주의적 접근
지형학에서 정량적 객관화와 실증주의적 접근을 이끈 대표적인 학자로 호톤(Horton)과 맥킨(Mackin), 스트랄러(Strahler) 등이 있다. Horton(1945)은 수리학적 개념을 바탕으로 유역 경관과 사면 침식, 지표 유출수를 정량적으로 설명하기 위한 틀을 마련하였다. 대표적으로, 하천 경관을 해석하는 데 하천 차수 개념을 제시하여 유역 네트워크 구조를 설명하였다. Mackin(1948)은 지형 요인이 하천 운반 퇴적물의 양과 유량 변동에 미치는 영향을 개념적으로 제시하여 수리기하학의 발달에 기여하였다. 호톤과 맥킨의 연구는 지형 프로세스를 수치화하여 표현하는 가능성을 열었고, 스트랄러에 이르러 ‘지형학의 동적 기반(dynamic basis of geomorphology)’ 개념으로 발전할 수 있었다. Strahler(1952)는 정량적이고 동적인 지형학이 응력(stress)의 구분, 물질의 구분, 변형(strain)의 구분을 기초로 함을 설명하였다. 그는 물리학, 생물학, 화학, 수학, 토양 역학, 수리학 등 타학문의 지식을 지형 경관을 이해하는 데 활용하였으며, t-검정을 도입하여 표본 평균 차이의 유의성을 현장에서 처음으로 검증하기도 하였다. 결론적으로, 정성적・서술적 지형학에서 정량적이며 프로세스(과정) 지향적인 동적 지형학으로 넘어가는 기초가 마련되었고 경관 연구에 근본적인 변화가 일어났다. 지형 형태와 프로세스를 통계 기법과 수학 모델링을 통해 정량적으로 분석하게 되면서, 보다 객관적인 설명이 가능해졌다. 이러한 과정은 타 학문의 패러다임을 수용하는 것으로 시작되었고, 자연스럽게 각 학문에서 시도되었던 정량적 접근법 역시 도입되었다. 그 결과, 개념적 차원에서의 논의를 넘어 지형 형성 과정(역학)에 초점을 맞춘 연구를 수행할 수 있었다. 아울러, 일부 지형학적 현상의 예측과 응용까지 가능하게 되었다(Morisawa, 1988). 그러므로 호톤과 맥킨으로부터 시작되어 스트랄러에 의해 극대화된 지형학의 정량적 객관화 흐름은 학제 간 연결과 융합을 통해 복합적인 분석을 가능케 한 예시가 된다. 이러한 학술적 분위기는 향후 지형학과 생태학, 진화생물학을 통합하고자 하는 생물지형학의 태동에 시발점이 되었다.
표준화된 기법을 개발하여 토양의 화학적 성질을 수치화하고, 토양층의 두께, 점토 함량, 풍화비 등의 물리적 속성을 측정하여, 여러 지역의 토양을 정량적으로 비교하였다. 루에(Ruhe)는 이러한 방법을 활용하여 특정 토양과 토양층의 발달이 지질 연대와 더불어 지형 기복과도 연관되었음을 밝혔다(Ruhe, 1956). 그의 연구지역인 미국 아이오와주 그린필드 인근에는 과거 빙기 및 간빙기에 형성된 다섯 가지 주요 지형 경관이 존재한다. 각 경관의 토양을 실트 비율 및 중광물/경광물의 풍화비로 비교한 결과, 특정 토양 혹은 토양층이 개별 지형 기복과 밀접한 연관을 보였으며, 토양의 유형 차이는 과거 고토양의 생성 연대별 특성을 반영하고 있었다. 종합하면, 루에의 논문은 지형학과 토양학을 연계했을 뿐 아니라 시공간적 범주를 넓힌 다중 관점의 고전으로 여겨진다. 한편, 토양 형성에 영향을 미치는 다양한 요인을 정량적으로 고려하는 시도는 ‘상태 요인 방정식(state factor equation)’을 제안한 예니(Jenny)의 연구에 잘 드러난다(Jenny, 1941). 그는 생태계의 속성과 토양 속성(s)이 기후(cl), 생물(o), 지형(r), 암석(p), 시간(t), 그리고 기타 요인(...)들과 가지는 긴밀한 관계를 수식으로 표현하여 토양 형성 프로세스를 설명하였다: s = f(cl, o, r, p, t, ...). 또한, 각 상태 요인의 영향력에 따라 그 기울기를 다르게 하여 변화율도 표현하였다. Huggett(1975)은 열린 시스템인 계곡 유역(valley basin)을 연구의 기본 단위로 설정하는 ‘토양 경관 시스템(soil landscape system)’ 개념을 제안하여, 토양 시스템의 구성 요소와 삼차원 유역 시스템의 경계를 정의하였다. 그가 시도한 토양 형성 삼차원 시뮬레이션은 이차원 토양층 중심의 기존 연구를 넘어 유역 분지 및 슈뢰브(Shreve)의 하천차수 개념과 연관된다. 실질적 지구 시스템에 가까운 방식으로 토양 형성 과정을 모델링했다는 측면에서, 그의 연구는 새로운 시공간적 관점을 제시한 연구로 볼 수 있다.
생태학자인 Curtis and McIntosh(1950)와 Whittaker(1967)는 구배 분석을 통해 경관 내 식생의 구조와 변이를 환경 요소, 각 식물종의 개체수, 군집의 특징 측면에서 파악하고, 식생의 공간적 분포를 설명하였다. 이들은 다양한 시스템의 식생과 환경 조건을 수치화하여 식생을 분류하는 데 그친 기존 생태학의 초점을 식물종 사이의 양적 관계를 밝히는 것으로 바꾸었다. 이를 통해, 식생과 환경 조건을 초공간(hyperspace)에 배열(ordination)하여 둘 사이의 관계와 그 구조를 정량적으로 표현할 수 있었다. 아울러, 추상적인 단위로 파악되던 식생을 복합적인 연속체로 인식할 수 있게 되었다. 한편, 휘태커(Whittaker)의 연구에서 생태학과 진화생물학의 만남을 암시한 구절을 찾아볼 수 있다. 그에 따르면 종 개체군의 분포는 일반적으로 환경구배를 x축으로, 개체수를 y축으로 하는 이항 곡선(binomial curve) 형태로 표현할 수 있다. 각 개체군 별 곡선의 형태와 최적점의 위치는 부분적으로 적응(adaptation)의 중심점과 유전적 분화의 범위를 나타낼 수 있다. 서로 다른 개체군 최적점의 위치, 식생 군집 내 지위(niche), 그리고 이로 인한 개체군 분포의 중첩을 통해, 개별 식물종은 적응적 차이(adaptive differences)를 향해 진화할 수 있다(Whittaker, 1967). 휘태커의 이론은 Gleason(1926)이 개체주의적 관점에서 종별 상호작용을 확인한 것을 넘어, 식물종의 생태적 지위와 유전적 분화까지 포괄하였다. Whittaker(1967)는 환경구배를 토대로 진화의 가능성까지 논의한다는 점에서, 현재 생물지형학의 전신과도 같은 위치를 점한다고 사료된다.
정량적 객관화와 실증주의적 접근을 통해 실세계의 다양한 요인을 수량화함으로써, 자연 현상을 통계적 기법과 수치모델을 활용하여 파악할 수 있게 되었다. 이와 더불어, 지형학, 토양학, 생태학 각 분야에서는 기초과학의 개념(혹은 범례)을 활용하여 관심 현상을 표현하기 시작하였다. 이러한 흐름은 시스템 안팎의 다양한 요소들 간 상호작용을 포괄하는 열린 시스템 이론을 바탕으로 확장될 수 있었다.
(4) 시스템 이론과 복잡계적 접근
자연과학에서 시스템 이론과 복잡계적 접근의 바탕이 되는 개념 중 하나는 탠슬리(Tansley)가 제시한 ‘생태계(ecosystem)’이다(Tansley, 1935). 그의 1935년 논문은 생태학을 기반으로 학제 간 통합과 융합이 구현된 대표적인 고전으로 여겨진다. 그는 클레멘츠와 필립스가 주장하던 기후에 의한 단방향(one-directional)의 점진적인 천이 이론(Clements, 1916, 1934; Phillips, 1931, 1934, 1935a, 1935b)에 반기를 들며, 교란에 의한 식생 파괴 및 새로운 식물종의 성장을 통해 퇴보하는 천이 혹은 역천이(retrogressive succession)의 가능성을 주장하였다. 또한, 토양, 지형, 방목, 화재, 벌목 등 다양한 요인에 의한 극상 형성을 설명하였고, 이 과정에서 생물군계(biome)와 물리적 환경 요소를 시스템적으로 인식할 수 있는 생태계 개념을 제시하였다(van der Valk, 2014). 기존의 생태학자들은 식생 군락을 유기체의 성장 과정에 빗대어 설명하며, 자연은 조화롭고 안정적인 상태로 향한다는 발달 은유적인 관점을 취했다. 반면, 탠슬리는 식생 군락을 온전한 유기체가 아닌 준유기체(quasi-organism)적 상태로 바라보아야 한다고 주장하였다. 식생 군락과 군집은 일부 유기체적 속성을 가지고 있으나, 스스로 번식하거나 안정성을 유지하는 실질적인 특성을 지니고 있지 않기 때문이다. 그의 주장은 변화하는 환경 조건에 따라 조절되는 시스템적 관점을 기반으로 자연을 설명하는 물리학 기반의 기계적 평형 은유로 평가된다(Tobey, 1981; van der Valk, 2014). 이는 20세기 후반 및 이후의 과학자들이 생태학을 바라보는 시각에 큰 영향을 주었다. 더 이상 자연을 균형과 안정 상태가 아닌, 요소들 사이의 상호작용을 토대로 하는 동적 평형 혹은 비평형적 관점으로 바라보게 된 것이다. 이러한 측면에서 생태계가 생물지형학의 태동기와 현재를 관통하는 핵심 개념이자 연구 대상이 된 것은 분명한 사실이다.
또 다른 생태학자인 왓(Watt)의 1947년 논문은 ‘스스로를 유지하고 재생하는 메커니즘’과 관련된 기념비적 논문이자 생태학적 고전으로 여겨진다. 식생 군집의 특성과 그 공간적 관계는 패치(patch) 구성의 총체적 관점에서 시간적으로 설명할 수 있다. 식생 군집의 변화는 군집 내 패치를 구성하는 종별 생애주기, 토양, 미지형과 미기후 변화 등을 반영하기 때문이다(Watt, 1947). 스승이기도 한 탠슬리의 생태계 개념을 이어받아, 왓은 시스템적 사고를 바탕으로 토양 단면과 기복 특성 분석에도 관심을 가졌다(van der Maarel, 1996). 그는 자연현상이 생태계를 기반으로 발생하기 때문에, 식생 군집과 관련 요소들을 융합하여 바라보는 것이 중요하다고 주장하였다. 왓의 논문은 1989년을 기준으로 생태학계에서 가장 많은 인용 횟수를 보였는데(McIntosh, 1989), 이는 향후 패치 역학(patch dynamics)으로 이어지는 초기 개념은 물론, 다양한 시공간적 범주를 고려한 시스템적 접근도 제시하였기 때문이다. 또한, “개별 식물은 환경의 일부 산물이며 그 자체가 환경을 변화시킨다.”라는 구절에서 오늘날 생물지형학과의 연결점을 찾을 수 있다.
시스템 이론의 발달은 20세기 중반 이후 평형 개념과 함께 시작되었지만(Renwick, 1992), 사실 자연에서의 평형에 대한 논의는 그 이전부터 이어져 왔다. 전술하였듯, 과거 학자들은 실세계가 균형이나 지향점에 이른 것을 자연스러운 상태로 생각하였다. 이는 지형학, 토양학, 생태학계에서도 주된 관점이었으며, 각 분야는 이를 바탕으로 발전하였다. 먼저, 데이비스의 연구에서는 침식과 퇴적이 균형을 이룬 상태인 평형하천(graded river) 개념이 제시되었다(Mackin, 1948). 지형학 전반에 걸쳐서는 두 가지 경우를 평형 상태로 설명했다. 하나는 특정 프로세스가 특정 지형 경관으로 귀결되는 것이고, 다른 하나는 퇴적물의 유출입 사이에 안정적이고 일관된 관계가 존재하는 것이다. 토양학에서의 평형 개념도 지형학과 유사하게 사용되었다. 토양학자들은 삭박에 의한 지표 물질의 제거와 기반암 풍화로 인해 생성되는 신선한 토양이 균형을 이루는 상황, 즉 침식과 토양 형성 사이의 균형을 평형으로 바라보았다(Phillips, 2010). 생태학에서는 클레멘츠의 극상 개념이 비슷한 맥락이다. 식생 군집은 기후의 영향을 받아 단계적인 순서를 통해 극상에 이르게 된다. 과거 생태학자들은 안정적이며 자가 유지가 가능한 극상을 정상적인 자연 조건으로 받아들였다.
전술하였듯, von Bertalanffy(1950)는 닫힌 시스템에서는 이러한 평형 상태에 도달하는 것이 가능함을 설명하였다. 그는 닫힌 시스템과 반대되는 개념으로, 시스템 내외의 상호작용을 강조하는 열린 시스템 개념도 제시하였다. 열린 시스템의 구성 요소들은 내부적으로 상호작용하는 동시에 외부 환경과 물질 및 에너지를 활발히 교환한다. 이후, 과학자들은 열린 시스템 이론을 통해 자연 현상을 다변량화하여 인지할 수 있었고(Chorley, 1962), 진정한 평형 상태가 존재하는지에 대한 답을 찾아가기 시작했다. 이 과정에서 평형 상태에 대한 반례가 꾸준히 확인되었고(Magilligan, 1992; Phillips, 1992, 2010), 교란에 의한 시스템의 역동성을 자연스러운 현상으로 인식하게 되었다(Sprugel, 1991). 결과적으로, 열린 시스템에서는 평형 상태가 존재하기 어렵다는 자연 경관의 비평형 패러다임이 발전하게 되었다.
보다 세부적으로, 필립스는 지형학과 토양학 분야에서 기존의 평형을 넘어선 비평형, 다중 평형 및 비선형성을 다루었다. 그가 제안한 ‘평형의 종말’은 단순히 전통적인 평형 개념을 거부하는 것이 아니다. 오히려 지표 시스템의 복잡하고 비선형적인 특성을 인정함으로써, 더욱 확장된 이론을 기반으로 지형학 연구를 수행함을 의미한다. Phillips (1992)는 평형 및 비평형 상태와 관련된 경관 요소가 동시에 존재하고, 각 현상은 동등하게 중요하다고 주장하였다. 즉 주어진 환경 제어 요인에 대해 여러 가지 평형 상태가 가능하며 시스템 또한 다양한 평형 상태를 지향하는 지형적 진화 궤적을 따를 수 있다고 설명하였다. 또한 미시적 규모에서의 평형을 불안정하고 일시적이라고 인식하는 것에 대한 대안이 필요함을 강조하였다.
한편, Phillips(2010)는 토양학의 ‘정상 상태 토양 두께(steady-state soil thickness; SSST)’ 개념이 실제 자연에서 보편적으로 적용되기 어렵다는 점을 지적하였다. SSST는 침식과 풍화가 동적 평형 상태를 이루어 일정한 토양 두께가 유지되는 것을 의미하며, 경관 진화 모델링이나 우주선 방사성 동위원소를 이용한 침식률・풍화율 추정에 유용한 기준으로 활용될 수 있다. 그러나 심층풍화와 레골리스(regolith) 제거, 침식과 풍화 사이의 비선형적 되먹임 등 다양한 요인으로 인해, 토양 두께는 항상 동적이며 균형 상태를 유지하기 어렵다. 따라서 SSST 개념은 특정한 환경에서 유용한 분석 틀로 작용할 수 있으나, 실제 토양과 풍화층의 복잡한 진화를 완전히 설명하기에는 한계가 있다. 지형학과 토양학 전반에 걸쳐 다양한 유형의 비평형과 비선형성을 다룬 필립스는 생물지형학적 관점으로 수렴하는 융합론적 접근을 보여주었다고 평가된다.
생태학 분야에서 Sprugel(1991)은 교란의 공간적 범위, 과거의 사건, 기후 변동성을 근거로 식생의 비평형을 다루었다. 초기 학자들에게 ‘자연적인’ 생태계는 대규모 교란(화재, 폭풍, 해일 등)이 없는 상태로 오랜 시간이 지난 후에 도달하는 공동체를 의미하였다. 그러나 현실적으로 거의 모든 생태계는 외부 교란의 영향을 받으며 변하기 때문에, 진정한 의미에서 자연적이고 안정적인 식생 구조를 정의하는 것은 매우 어렵다. 우연한 요인이나 작은 기후변화가 식생에 매우 큰 변화를 불러올 수 있으며, 이로 인해 특정 경관의 생물상 및 생태계 프로세스는 상당한 기간에 걸쳐 크게 달라질 수 있다. 스프루겔(Sprugel)이 제시한 식생 군집과 천이의 불안정성 혹은 비평형성은 극상에 대해 다시 생각해보는 계기가 되었다. 특정 시점의 환경 변화가 식생 구조에 미치는 중장기적인 영향은 예측하기 어렵기 때문에, 근본적으로 진정한 자연 식생이 존재하는 것인지에 대한 답을 찾는 것은 불가능하다. 이런 측면에서, 스프루겔은 생태학의 비평형 및 비선형적 요소를 다룸으로써 오늘날 생물지형학의 비선형성 이론에 지대한 영향을 주었다.
마지막으로, 비평형과 관련된 논의는 복잡계적 사고로 연결되었다. 기존의 시스템 이론을 바탕으로, 임계치, 되먹임, 결정론적 혼돈, 비선형성 등을 고려하여 비평형 상태의 경관을 설명하려는 시도가 이어졌다. 특히, 에너지와 물질의 산출이 투입된 양에 비례하지 않는 시스템의 비선형성은 시스템 내 구성 요소 간 상호작용을 바탕으로 특정 현상이 증폭되거나 감쇄될 수 있음을 보여주었다. 복잡계적 사고는 실세계의 복잡한 현상을 표현하기 위해 교란과 시스템 내외의 상호작용을 포함하는 개념적 틀로서 지금도 학문적 논의의 범위를 넓혀가고 있다(Kim, 2017; Stallins and Parker, 2003). 대표적으로, Stallins(2006)는 복잡계와 비선형 동적 시스템(nonlinear dynamical systems)으로의 패러다임 전환을 통해 생물지형학의 태동을 설명할 수 있다고 주장하였다. 그는 학계를 아우르는 비평형성 및 비선형적 관점이 복잡계 이론에서 제시하는 ‘초기 조건의 변화’와 연결될 수 있다고 믿었다. 이에, 복잡계 적응 시스템(complex adaptive systems; CAS) 이론을 도입하여 해안사구의 식생과 지형이 재귀적 자기 조직화(recursive self-organization)를 이루는 과정을 탐색하였다(Stallins and Parker, 2003). CAS에서 재귀적 자기 조직화는 경관의 진화에 수렴이나 발산과 같은 방향성을 부여할 수 있다. 이러한 방향성은 클레멘츠가 주장하던 결정론적 방향으로 천이를 제한하는 것이 아니라, 오히려 결정론과 우연성을 포괄하며 평형, 다중 평형, 비평형 관점을 통합한다(Stallins and Parker, 2003). 결론적으로, 스탈린스는 해안사구와 같이 반응성이 높은 지표 시스템에서는 지형과 퇴적물의 이동, 식생을 독립적인 요소들로 간주할 수 없음을 역설함으로써 생물지형학이라는 새로운 복합과학의 장을 마련한 융합론자로 사료된다(Kim and Yu, 2009).
3) 생물지형학의 발달사 조망: 주요 학술행사 소개
본 소절에서는 1980년대 후반부터 시작된 생물지형학 관련 학술대회들의 서문(preface)을 토대로 생물지형학의 발달사를 파악한다. 이를 바탕으로 생물지형학의 현재와 미래에 대한 시사점을 제공하고자 한다. 대표적으로 북아메리카 대륙의 지형학 관련 학술행사인 빙햄턴 지형학 심포지엄(Binghamton Geomorphology Symposium)과 미국 지구물리학회(American Geophysical Union)를 소개한다.
1970년부터 시작된 빙햄턴 심포지엄은 생물지형학을 주제로 오늘날까지 총 다섯 번의 연례 행사를 개최하였다(Baker, 2020; Viles, 2020). 그 첫 번째는 1995년 제26차 심포지엄으로, 주제는 ‘Biogeomorphology, terrestrial & freshwater systems’였다. Hupp et al.(1995)의 심포지엄 서문은 생물지형학을 생물상과 지형 형태 및 과정 사이의 관계를 연구하는 학문(Viles, 1988)으로 정의하며, 코울스를 비롯한 초기 생태학자들이 종의 분포를 설명하기 위해 지표 형태를 기술할 필요성을 느꼈다고 언급하였다. 특히, 지형학의 길버트와 데이비스, 생태학의 코울스, 클레멘츠, 글리슨 등 초기 이론가들이 동시대를 공유하며 서로 근본적인 영향을 미쳤음을 논하였다. 그럼에도 불구하고 학제 간 연구가 20세기 후반까지 거의 이루어지지 않아 생물지형학이 여전히 초기 발전 단계에 머물러 있음을 강조하였다. 해당 심포지엄은 본래 식생과 지형의 상호작용에 중점을 두고자 했으나, 동물과 지형의 상호작용에 대한 관심이 커짐에 따라 생물지형학 전반을 다루는 방향으로 확장되었다(Hupp et al., 1995; Viles, 2020). 또한, 지형학과 생태학의 연결을 처음으로 시도한 것이 Hack and Goodlett(1960)임을 언급하며, 이 연구를 검토하고 재평가하는 것으로 심포지엄의 문을 열었다.
두 번째로, 2005년 제36차 심포지엄에서는 ‘Geomorphology & ecosystems’를 주제로 이전보다 폭넓은 생물지형학적 논의를 시도하였다. 해당 심포지엄은 생태계 내에서 지형학의 역할, 지형 시스템 내에서 생태학의 역할, 지형 생태계의 통합적 관리를 다루는 세 가지 소주제로 구성되었다. Renschler et al.(2007)는 서문에서, 새롭게 등장한 환경 문제의 해결을 위해 학제 간 경계가 흐려지면서 지형학과 생태계 생태학(ecosystem ecology)의 결합을 통해 환경 과학의 새로운 패러다임이 형성되었음을 밝혔다. 아울러, 지형학과 생태학이 각자 평행한 발전 과정을 거쳐왔음에도 불구하고 통합적이며 포괄적인 이론을 발전시키지 못했고, 필요한 경우에만 다른 분야의 정보나 이론의 일부를 취사선택하였음을 언급하였다. 이는 지형학과 생태계 생태학 분야에서 물리적 경관을 고려하는 방식에 근본적인 차이가 존재하는 것과 연관된다. 따라서, 2005년 빙햄턴 심포지엄의 목표는 지형학과 생태계 생태학 사이의 이론 및 응용 격차를 해소하는 것이었다. 이와 관련하여, 해당 심포지엄은 경계, 개념적 틀, 비선형적 반응 등 지형 생태계(geomorphic ecosystems)에 대한 학제 간 연구의 주요 과제를 다루었다(Renschler et al., 2007; Viles, 2020).
생물지형학과 관련된 세 번째 빙햄턴 심포지엄은 2009년 제40차 ‘Geomorphology & vegetation: interactions, dependencies & loops’였다. 해당 심포지엄은 1995년 행사를 재검토하면서 특히 식생과 지형 시스템 사이의 연관성에 중점을 두었다(Hession et al., 2010). 서문에 따르면, 지형학에서 식생의 중요성이 강조됨에 따라 연구의 복잡성이 훨씬 높아졌고, 이는 생물이 지표 프로세스에 미치는 영향을 이해하는 데 어려움을 더했다(Dietrich, 2008). Hession et al.(2010)은 이러한 어려움이 증폭되어, 생물지형학이 ‘잠재적으로 상호보완적이지만 대체로 교차하지 않는 연구 분야’로 이어지는 것을 경계했다. 따라서 생명과 경관이 불가분의 관계에 있음을 인정하며(Reinhardt et al., 2010), 둘 사이의 역동적인 상호작용을 이해하기 위한 ‘퍼즐 조각’을 모으는 것이 해당 심포지엄의 목표임을 강조하였다. 이를 위해 공학자, 지구과학자, 생태학자가 모여 다양한 연구 방법을 통해 복잡한 시스템에 대한 이해를 높여야 한다고 주장했다. 이러한 맥락에서 유역 구조를 중심으로 하는 다양한 시스템에서의 식생과 지형 형태・프로세스 사이 되먹임을 다루는 연구들이 발표되었다(Hession et al., 2010; Viles, 2020).
네 번째로 개최된 생물지형학 중심의 빙햄턴 심포지엄은 2011년 제42차 ‘Zoogeomorphology & ecosystem engineering’이었다. 해당 심포지엄의 목적은 지형학적 접근법과 생태학적 접근법 사이의 간극을 좁히고, 동물지형학적 연구와 생태계 엔지니어링 연구를 연결하는 것이었다(Butler and Sawyer, 2012). 동물이 가지는 지형학적 중요성은 일찍이 다윈의 지렁이 활동과 분변토 형성에 대한 연구를 통해 언급되었지만(Darwin, 1881), 생태학과 지형학 사이의 상호작용은 20세기 후반까지도 지형학 문헌에서 크게 주목받지 못하였다(Butler and Sawyer, 2012; Stine and Butler, 2011). Butler and Sawyer(2012)는 1980년대 후반 바일스의 저서(Viles, 1988)를 통해 생물지형학이 대두하며 지형과 생태 사이의 상호작용에 대한 연구가 ‘부활’했음에도 불구하고, 지형학과 생태학은 평행선을 달리며 통합을 이루지 못했다고 지적하였다. 동물지형학과 생태계 엔지니어링 개념 역시 비슷한 주제를 다룸에도 불구하고 서로 독립적으로 발전해왔기에 두 학계 사이에 상호작용이나 시너지 효과가 부족했음을 강조하였다. 이러한 입장을 기반으로, 해당 심포지엄은 미생물과 수권 군집, 대형 동물과 생태계 엔지니어링, 퇴적물 이동과 침식에 영향을 미치는 다양한 동물 등 세 가지 소주제로 구성되었다(Viles, 2020).
가장 최근에 개최된 생물지형학 관련 빙햄턴 심포지엄은 2017년 제48차 ‘Resilience and bio-geomorphic systems’였다. 해당 심포지엄은 사회생태학적 회복탄력성 사고(resilience thinking)와 생물지형학, 지형학 간의 연계성을 명확히 하고 그 간극을 좁히는 것을 목표로 하였다. 이를 통해, 지금까지 언급한 다섯 개의 심포지엄 중 생물지형학을 가장 폭넓게 다루었다(Thoms et al., 2018; Viles, 2020). Thoms et al.(2018)의 서문에 의하면, 지형 프로세스는 생태 및 사회 시스템과 병행하여 다양한 시공간적 규모에서 작동하고 서로 총체적인 영향을 미친다. 따라서, 생물지형학적 접근법은 되먹임, 상호작용, 임계치, 반응 등의 개념을 통합하여 지형학과 사회생태 시스템을 연결하는 경로를 만들 수 있다(Thoms et al., 2018). 이와 관련하여, 해당 심포지엄은 회복탄력성 사고를 기반으로 지형 시스템, 극한 현상과 임계치, 생태와 지리의 연결, 동물지형학, 연안 지형학, 사회-생태 시스템 등 여섯 개의 소주제로 구성되었다.
빙햄턴 지형학 심포지엄 외에도 미국 지구물리학회의 2008년 가을 연례학술대회 역시 ‘Multiscale feedbacks in ecogeomorphology’를 주제로 하는 분과를 통해 생물지형학을 다루었다. 지형 시스템은 물리적-생물학적 되먹임을 통해 비선형적 특성을 가지기 때문에, 경관 변화와 관련된 시스템의 반응을 이해하거나 예측하는 것이 매우 어렵다(Wheaton et al., 2011). 그럼에도 불구하고, 최근 생태계 건강성(ecosystem health)에 대한 우려가 커지면서 이러한 되먹임 작용을 규명하려는 연구가 활발하게 진행되고 있다(Chaffin and Scown, 2018; Darby, 2010; National Research Council, 2010; Rice et al., 2012; Viles et al., 2008; Wheaton et al., 2011). 해당 분과는 습지와 하구, 강, 고지대에 이르는 다양한 환경에서 생물학적 되먹임과 지형학적 되먹임 사이의 대조적인 관점을 통합함을 목표로 하였다.
상술하였듯, Viles(2020)는 지난 30여년 동안 다양한 학술행사가 진행되며 생물지형학의 이론이 세 가지 측면에서 점진적으로 발전하였다고 보았다. 첫째, 개별 종과 생물지형학적 상호작용을 다루는 정성적 연구에서 복잡한 상호 의존성을 검증하는 정량적 연구로의 전환이 이루어지고 있다. 둘째, 생물지형학적 연구의 간학문적 수준과 다양성이 향상되었다. 셋째, 전지구적 환경 변화의 광범위한 틀에서 생물지형학 이론이 개발되고 있다. 그럼에도 불구하고, 2000년대 이전까지는 지형학계에서 생물지형학적 상호작용을 진지하게 받아들이지 않았던 것도 사실이다(Viles, 2020).
결론적으로, 생물지형학의 현재와 미래에 대해 두 가지의 시사점을 유추할 수 있다. 첫째, 공통된 연구 질문과 패러다임을 중심으로 진정한 학제 간 연결 및 융합이 지속적으로 요구된다. 최근의 학술대회에서도 생물지형학과 연관 분야 사이에 긴밀한 소통과 실질적인 공동 연구가 부족하다는 인식이 있었다(Butler and Sawyer, 2012; Hession et al., 2010; Hupp et al., 1995; Renschler et al., 2007). 그 원인으로는 학문 분야 사이에 존재하는 패러다임, 연구 방법론, 연구 스케일 차이가 있으며 이는 단기간에 해결하기 어려운 문제이다. 둘째, 생물-지형 복합체가 보이는 비선형성과 복잡성을 이해하고 적용하는 것이 중요하다. 생물지형학은 지형학과 생태학, 더 나아가 진화생물학까지 아우르는 복합학문으로, 다양한 구성 요소들의 비선형적 상호작용을 탐색한다(김대현, 2021; Hession et al., 2010; Renschler et al., 2007; Thoms et al., 2018; Viles, 2020; Wheaton et al., 2011). 따라서 지형과 생물 사이의 되먹임을 이해하고 외부 조건의 변화에 대한 시스템의 반응을 예측하기 위해서는 다양한 시공간적 스케일을 고려할 필요가 있다(Kim and Zheng, 2011; Kim et al., 2012).
4) 자연지리학과 생물지형학의 연결 고리
자연지리학과 생물지형학은 서로 긴밀하게 상호작용하며 유사한 발달 과정을 보여왔다. 두 학문은 모두 자연에서 발생하는 다양한 현상의 과정과 결과를 토대로 지표 시스템을 이해하는 데 중점을 둔다. 자연지리학은 지형, 기후, 생태, 수리・수문 등 지구의 다양한 요소가 보이는 공간적 변이를 탐구한다(Davis and Snyder, 1898; Strahler and Strahler, 2003). 생물지형학은 생물과 지형이 하나로 연결된 복합체라는 관점을 바탕으로 이들 간의 상호작용과 되먹임을 연구하는 종합학문이다(김대현, 2021; 김대현 등, 2020; Viles, 1988). 자연지리학과 생물지형학은 큰 틀에서 지표 시스템을 연구한다는 공통점을 가지고 있지만, 그 연결점이 무엇인지는 분명하지 않다. 이 연결고리를 정의할 수 있다면, 복잡한 지표 시스템에 대한 이해를 확장할 수 있을 것이다. 또한, 이건학 등(2024)이 논의한 생물지형학적 되먹임 연구에 따르는 난점(공간적 스케일, 시간적 스케일, 생물 종별 차이, 개체 생애 주기별 차이, 서식지 특성의 문제)을 해결하는 데에도 기여할 수 있다. 본 논문에서는 두 학문 간 연결고리를 보여주는 사례로 하천생물지형학적 천이(fluvial biogeomorphic succession; FBS)를 소개한다. FBS는 하천 지형과 식생 군집 사이의 생물지형학적 구조와 프로세스가 선택 및 재생산되는 하천 경관 단위의 순차적 발달 과정을 의미한다(Corenblit et al., 2007). 본 소절에서는 하천생물지형학적 상호작용과 이를 바탕으로 제시된 FBS 개념이 자연지리학과 생물지형학을 어떻게 연결할 수 있는지 살펴본다.
하천생태계는 지형, 수리・수문, 식생으로 구성된다. 하천생물지형학적 상호작용은 이들이 서로 영향을 주고받는 복합적인 현상을 의미한다(Corenblit et al., 2007, 2011; Kim, 2012). 식생은 수리・수문과 지형 모두에 영향을 미치고, 이렇게 생기는 새로운 환경 조건은 다시 식생 군집에 영향을 미친다. 예를 들어, 식생은 하천 흐름에 저항으로 작용하여 유속을 감소시킨다. 또한 식생은 부유 물질을 포획하여 퇴적을 촉진시키고, 뿌리를 통해 지형의 안정성을 높여 침식을 줄일 수 있다. 반대로, 하천 지형은 수위, 침수 빈도, 퇴적물 특성, 영양분 공급 등 다양한 요소를 조절하여 식생 군집의 구성과 분포에 영향을 준다. 이렇듯 하천생태계의 구성 요소들은 서로 일방향적인 관계가 아니라 긴밀하게 상호작용하는 관계를 가진다. 이러한 순환 과정은 식생, 수리・수문, 지형 등 다양한 요인이 혼재된 하천생태계의 역동성을 나타낸다(김대현, 2021; Corenblit et al., 2007; Diehl et al., 2017). 하천생물지형학적 상호작용은 후술될 FBS를 이해하는 데 기반이 된다.
(1) 하천생물지형학적 천이(FBS)
FBS는 하천생태계의 주요 구성 요소 간 상호작용을 모델로 구축하기 위해 제안된 개념이다. 이 개념은 Holling and Gunderson(2002)의 적응 주기(adaptive cycle) 이론을 기반으로 하천생태계의 시스템 역학을 해석한다. 적응 주기 이론은 파괴와 재조직 과정을 통해 생태계가 변화에 적응하며 진화하는 동적 시스템임을 보여준다(Corenblit et al., 2007). FBS는 하천생물지형학적 상호작용을 중심으로 하천생태계 경관에서 나타나는 회복탄력성과 역동성을 설명한다.
하천생태계에서는 환경 조건의 변화에 따라 특정 요소의 영향이 상대적으로 우세하게 나타난다. FBS는 이를 지형적(geomorphic), 선구적(pioneer), 생물지형적(biogeomorphic), 생태적(ecological) 단계로 구분한다. 지형적 단계는 대규모 홍수와 같은 강한 교란이 발생한 직후에 나타나며, 하천 지형이 재형성되고 기존 식생이 완전히 파괴된다. 이 단계에서는 수리・수문과 지형 요소의 영향이 우세하게 작용하여 침식과 퇴적을 주도한다. 선구적 단계는 지형적 단계에서 형성된 나지에 선구종이 정착하고 성장하는 시기이다. 성장과 번식이 빠른 선구종은 하천 환경의 불안정한 조건에 쉽게 적응한다. 수리・수문과 지형 요소의 영향이 여전히 우세하지만, 점차 식생의 영향력도 증가한다. 선구종의 정착과 성장은 하천 흐름과 퇴적물 이동에 영향을 미치며 지형변화에 기여한다. 생물지형적 단계는 식생의 영향이 가장 큰 시기이다. 충분히 성장한 식생은 침식을 감소시키고 퇴적을 촉진함으로써 하천 제방을 안정화하고 사주와 범람원 등을 형성한다. 이 단계에서 식생 군집은 더욱 복잡해지며, 식생, 수리・수문, 지형요소 사이의 균형이 형성되기 시작한다. 생태적 단계는 수리・수문 및 지형적 교란의 영향이 줄어드는 시기이다. 이 단계에서는 경쟁과 천이와 같은 생태적 프로세스가 우세해지며 식생 군집은 하천 지형을 유지하고 형성하는 데 중요한 역할을 한다. 생태적 단계는 큰 교란이 없는 한 오랫동안 지속될 수 있다(Corenblit et al., 2007).
FBS는 하천생태계를 구성하는 여러 요소들이 되먹임 작용을 통해 서로 영향을 주고받으며 복잡하게 얽혀 있음을 나타낸다. 지형적 단계는 홍수와 같은 교란이 하천 지형을 재형성하고 생태계를 초기화하여 선구종에게 새로운 정착 기회를 제공한다. 지형적 단계에서는 선구 단계의 식생 발달을 위한 토대가 마련되는 것이다. 선구적 단계에서는 식생, 수리・수문, 지형 요소 모두가 상호작용하기 시작하며, 시간이 지나며 각 요소가 더욱 긴밀한 관계로 발전하게 된다. 생물지형적 단계에서는 식생이 수리・수문과 지형의 변화를 주도하며, 식생에 의한 환경 변화가 다시 식생 군집의 발달과 천이에 영향을 미친다. 이는 하천생태계의 역동성을 보여주는 핵심적인 단계이다. 마지막으로 생태적 단계에서는 하천생물지형학적 상호작용이 줄어들고 생태적 프로세스가 우세해진다. FBS는 네 가지 주요 단계로 나뉘어 있지만 각 단계는 명확히 구분되지 않으며, 강한 교란이 발생하면 어느 단계로든 되돌아갈 수 있다(김대현 등, 2020; Corenblit et al., 2007, 2009; Kim and Kupfer, 2016).
(2) 자연지리학과 생물지형학의 연결
FBS는 자연지리학과 생물지형학의 교차점에서 중요한 연결고리가 될 수 있다. 앞서 언급했듯, 생물지형학이 탐구하는 경관의 생성과 변화는 지구의 물리・화학적 특성, 지형, 기후, 식생, 수리・수문적 현상 등을 분석하는 자연지리학의 주된 주제이기도 하다. 이 두 학문은 경관 변화와 생태계의 상호작용을 이해하려는 공통된 목표를 가지며, FBS는 이를 통합적으로 설명할 수 있는 틀을 제공한다. 특히, FBS는 두 학문에서 공통적으로 다루는 역사적 우연성, 비선형성, 다중 경로와 같은 핵심 개념을 포함하고 있다. 역사적 우연성, 비선형성, 그리고 다중 경로 개념은 예측 불가능하고 복잡한 자연 시스템의 변화 과정을 설명할 수 있다는 점에서 두 학문의 연결 지점을 강화한다(Corenblit et al., 2007; Phillips, 1999b; Stallins, 2006). 즉, FBS는 복잡계로서 자연 환경을 분석하는 기반으로, 자연지리학과 생물지형학의 연결고리로 기능한다.
역사적 우연성은 초기 조건의 차이가 장기적으로 자연 시스템의 발전 경로에 중대한 영향을 미친다고 강조한다. 이는 특정 사건이나 초기 조건이 단순히 과거의 흔적으로만 남는 것이 아니라, 시스템의 현재 상태와 미래의 변화 방향에도 영향을 미칠 수 있음을 의미한다(Stallins, 2006). 비선형성은 자연 시스템이 선형적 인과관계를 따르지 않으며, 시스템 내 상호작용에 따라 임계치를 넘어가면 급격히 변할 수 있음을 설명한다. 시스템의 아주 작은 변화가 예상보다 큰 영향을 미쳐 특정 조건을 초과하면, 갑작스럽고 극적인 전환으로 이어질 수 있다(Brunsden and Thornes, 1979; Holling, 1973). 다중 경로는 동일한 초기 조건에서도 시스템이 상이한 경로를 통해 전혀 다른 결과에 도달할 수 있음을 설명한다. 시스템 외부 요인의 영향과 내부 요소 간 상호작용을 강조하여 같은 환경 조건이 다양한 결과로 이어질 수 있는 자연 시스템의 복잡성을 드러낸다(Corenblit et al., 2007; Phillips, 1999a). 이처럼 역사적 우연성, 비선형성, 다중 경로는 자연 시스템이 어떻게 작동하고 변화하는지에 대해 복잡계적 사고를 기반으로 한 통합적 이해를 제공한다.
(3) 자연지리학과 생물지형학에서 나타나는 역사적 우연성, 비선형성, 다중 경로
경관의 생성과 변화는 자연지리학과 생물지형학 모두에게 중요한 주제이다. 또한 지구의 물리・화학적 특성, 지형, 기후, 식생, 수리・수문적 현상 등은 두 학문 공통의 연구 대상이다. 두 학문은 경관 변화와 생태계의 상호작용을 이해하려는 공통된 목표를 가지며, FBS를 통해 두 학문의 공통된 핵심 개념을 통합적으로 설명할 수 있다. 따라서 본 소절에서는 하천생태계를 예시로 역사적 우연성, 비선형성, 다중 경로가 어떻게 다뤄지는지 살펴보고, FBS가 두 학문의 연결고리가 될 수 있는 이유를 논하고자 한다.
자연지리학에서는 위 세 가지 핵심 개념이 경관을 분석하는 데 중요한 역할을 한다. 역사적 우연성은 초기 조건이나 특정 사건이 이후의 경관 변화 및 경관 형성 과정에서 결정적인 역할을 하는 것을 설명한다. 예를 들어, 강우가 증가하면서 특정 지역에서 홍수가 발생하여 많은 퇴적물이 이동하기 시작했다고 하자. 홍수로 인한 퇴적물의 이동과 분포는 기존 하도의 확장 또는 새로운 하천 네트워크의 형성이라는 결과를 초래할 수 있다. 수리・수문 패턴 변화와 같은 특정 초기 조건은 여러 요소들과 복합적으로 작용하여 하천생태계의 장기적인 발전 경로를 형성할 수 있다. 비선형성은 동적이면서도 복잡한 경관 형성을 설명한다. 자연 시스템에서의 작은 변화는 예상치 못한 결과를 초래할 수 있다. 예를 들어, 하천의 유속이 미세하게 증가했음에도 특정 임계치를 초과하게 되면 하도의 침식이 급격히 가속화되어 하천의 방향이 바뀔 수 있다. 다중 경로는 동일한 조건에서도 다양한 발전 경로가 발현되어 전혀 다른 경관이 나타날 수 있음을 설명한다. 예를 들어, 홍수로 인해 하천이 깊어지고 침식이 가속화되는 경우가 있는 반면, 퇴적물이 쌓여 새로운 유로가 형성되는 경우가 있을 수 있다(Phillips, 2006, 2009).
생물지형학에서 역사적 우연성은 생태계와 지형 간 상호작용에 초점을 맞춘다. Stallins(2006)에 따르면, 특정 시점의 사건과 상호작용이 시간의 흐름에 따라 누적되고, 현재와 미래의 구조 및 기능에 영향을 미친다. 이는 단순한 인과관계를 넘어, 복잡한 피드백과 시간적 누적으로 경관과 생태계가 형성되고 유지됨을 강조한다. 예를 들어, 침수와 같은 교란이 빈번한 환경에 적응한 종에 의해 낮은 지형이 형성되어 이후 교란에 대한 저항성을 낮아지고, 교란이 드문 환경에서는 퇴적을 촉진하는 식물이 높은 지형을 만들어 교란에 의한 충격이 완화된다. 이러한 상호작용은 과거 교란이라는 특정 사건이 현재와 미래의 종 조성과 경관 시스템에 영향을 미침을 보여준다. 비선형성은 생태계와 지형이 서로 영향을 주고받으며 급격한 변화를 일으키는 특성을 설명한다. 작은 초기 변화라도 생태계와 지형 간의 상호작용 안에서 되먹임 순환을 통해 예상 밖의 결과를 초래할 수 있다(Phillips, 1999b). 예를 들어, 식생이 수리・수문 조건을 약간만 바꿔도, 시간이 지남에 따라 경관 내 기복을 크게 변화시키는 결과를 초래할 수 있다(Corenblit et al., 2011). 다중 경로는 초기 조건에 따라 생태계와 지형이 다양한 발전 경로를 가질 수 있음을 의미한다. 동일한 초기 조건을 가진 하천이라 하더라도, 외부 교란의 영향력과 상호작용 방식에 따라 발전 경로는 상이할 수 있다. 예를 들어, 홍수로 인해 식생이 제거되고 하상이 급격히 변할 경우, 해당 지역은 새로운 생물-지형 시스템으로 재구성될 수 있다. 반면, 동일한 홍수 상황에서도 식생이 침식으로부터 효과적으로 퇴적물을 보호할 경우, 하천생태계의 변화는 크지 않을 수 있다. 이렇게 다양한 발전 경로는 생물지형학에서 경관이 어떻게 형성되고 변화하는지 설명하는 데 중요한 역할을 한다(Corenblit et al., 2007).
경관의 변화를 설명하기 위해서는 생태계의 구성요소와 상호작용을 충분히 이해해야 한다. FBS는 자연지리학의 맥락에서 하천생태계의 다양한 구성요소를 포함하고, 생물지형학의 맥락에서 각 구성요소 간 복잡한 상호작용을 설명하는 개념이다. 이러한 측면에서 FBS는 자연지리학과 생물지형학을 잇는 연결고리로 기능하고, 더 나아가 복잡계로서의 하천 경관이 변화하는 과정을 통합적으로 이해하는 가능성을 제공한다.
5) 소결
두 가지 이상의 학문 분야를 통합하여 연구하는 것은 쉽지 않다. 이는 각 학문이 핵심적으로 다루는 연구 질문과 발전 방향이 서로 상이하기 때문이다. 특정 학문 분야의 핵심 질문은 해당 학계의 지배적인 연구 패러다임을 형성하며, 오랜 기간 동안 연구의 우선순위를 결정짓는다. 때에 따라, 서로 다른 학문 분야의 연구 질문이나 패러다임이 수렴할 경우 새로운 통찰력이 도출될 수 있다(Renschler et al., 2007). 생물지형학은 지형학, 생태학, 진화생물학이 통합적인 시각 아래 융합된 새로운 복합학문으로 여겨진다. 이러한 생물지형학의 통찰력을 파악하려면, 관련 분야의 발달사와 패러다임의 변화 과정을 깊이 이해해야 한다. 표 1에 제시된 연구 사조를 바탕으로, 지형학, 토양학, 생태학 분야 주요 학자들의 학문적 위치를 시간의 흐름에 따라 시각화한 결과는 그림 1과 같다. 학제 간 연구가 활발해지고 각 학문의 연구 방향이 통합론적 관점에 가까워질수록 생물지형학으로 수렴하는 경향이 나타남을 확인할 수 있다. 또한, 개별 학자들이 상호 영향을 주고 받거나 협업을 통해 생물지형학적 접근을 발전시키는 사례도 관찰된다. 대표적인 예시로, 평형론자 지형학자인 핵과 평형론자 식물 생태학자인 구들릿의 공동 논문은 애팔래치아 유역의 지형과 생태를 통합한 연구로(Hack and Goodlett, 1960), 생물지형학의 전신으로 여겨진다(Hupp et al., 1995; Osterkamp and Hupp, 1996).

정리하면, 지형학, 토양학, 생태학의 발전 과정은 생물지형학의 형성과 발전에 중요한 영향을 미쳤다. 각 분야는 독립적인 발전 과정 속에 지식과 방법론을 축적하며 자연환경의 복잡한 상호작용을 이해하는 데 중요한 기반을 제공하였다. 복합학문인 생물지형학은 간학문적인 접근을 통해 지구 시스템을 종합적으로 이해하고 다양한 환경 문제를 예측하고 대비하는 데 핵심적인 역할을 할 잠재력을 지닌다. 특히, 지형학, 토양학, 생태학의 융합을 통해 도출된 통찰과 방법론을 정량적 연구로 발전시킨다면, 급변하는 자연환경에 효과적으로 대응할 수 있을 것이다. 더 나아가, 각 학문 분야에서의 창의적인 접근과 연구 결과가 새로운 패러다임 전환을 일으켰듯이, 생물지형학에서도 진화생물학과의 만남을 통해 새로운 패러다임이 도래할 것으로 기대된다.
4. 자연지리학과 진화생물학의 만남
1) 생물지형학의 현재
본 논문의 문제의식은 생물지형학이라는 신생 학문 분야에서 비롯되었다(김대현, 2021; Kim and Lee, 2022; Kim et al., 2024). 기본적으로 지형학자는 중력, 물, 바람, 빙하와 같은 환경 인자가 지표면의 형태를 변화시키는 과정을 탐구한다. 이런 전통 속에서 식물과 동물, 미생물 등의 유기체가 지형학적 현상에 기여하는 역할은 큰 주목을 받지 못했다. 그러나 오늘날 학계는 지구상에서 발생하는 거의 모든 지형형성작용에 생물이 직간접적으로 영향을 주고 있다고 판단한다(Corenblit et al., 2007; Phillips, 2016; Viles, 2020). 암석의 풍화와 침식, 물질의 운반과 퇴적, 심지어 지반의 융기와 침강에 이르기까지, 땅의 형태를 변화시키는 모든 작용에 생물의 활동이 개입되어 있다는 것은 자연선택과 마찬가지로 가설의 수준을 넘어 보편적인 이론으로 받아들여지고 있다. 건조 지역에서 모래가 날리고 쌓이는 현상마저 지의류와 조류, 균류, 박테리아 등으로 구성된 생물 박피(biological crust)의 역할을 배제한 채 설명할 수 없음이 다수의 현장 연구를 통해 검증되고 있다(Ravi et al., 2010; Viles, 2012). 이런 문제의식 속에 1980년대 후반에 이르러 생물지형학이 정식 용어로 확립되고 독립된 분야로 인정받기 시작하였다(Viles, 1988, 2020).
본 논문의 맥락에서 생물지형학의 가치는 생물이 주변 환경을 적극적으로 변화시킴을, 다시 말해 지표 변화의 능동적 주체임을 학문의 전면에 내세웠다는 데 있다. 이는 기존의 생태학과 생물지리학, 진화생물학에서 생물의 생존과 분포, 생식, 다양성 등을 주변 환경 조건에 좌지우지되는 것으로, 즉 생물을 다분히 자연계의 수동적인 존재로 인식해 온 것과 대조적이다. 고도에 따라 기온, 습도, 토양의 성질이 다르기 때문에 형성되는 식생의 대상 분포(vegetation zonation)가 대표적인 예이다(Morueta-Holme et al., 2015; von Humboldt and Bonpland, 1807). 아울러, 자연선택은 용어 자체에서 생물종의 이러한 수동적인 위치가 뚜렷하게 드러난다.
물론, 이러한 전통적 관점이 문제시될 필요는 없다. 이미 다양한 자연과학 분야에서 생물을 - 통계적으로 말하면 - 종속변수로 설정하고 눈부신 학문적 발전을 이루어 낸 바 있다. 다만, 생물과 주변 환경 사이의 관계를 서로 능동적인 영향을 긴밀하게 주고받는 되먹임의 맥락에서 바라볼 때 새로운 관점을 발견할 수도 있다. 예를 들어, 기후변화로 인한 해안 침수가 예측되고 이것이 사회적 문제로 대두되는 상황을 일부 생물지형학자들은 식물의 역할을 무시한 데서 비롯된 ‘호들갑’이라고 주장한다(Kirwan et al., 2016). 해안습지에서 식물은 만조시 조류의 속도를 감소시키고 떠다니는 미세 입자를 포집함으로써 퇴적률을 높인다. 오랜 시간이 지나면서 습지 표면의 고도는 점진적으로 상승하여 침수 빈도가 감소하게 된다. 이는 곧 습지 식물의 스트레스가 감소하는 것이기에, 조만간 이 장소에서 더 무성한 식생 피복을 기대할 수 있다. 이는 결국 퇴적률과 고도의 추가적인 상승으로 이어질 것이다. 이렇게 해안습지는 꾸준히 성장하고 - 연간 10mm를 넘지 않는 수준이라면 - 해수면 상승에 자연적으로 대처할 수 있는 것이다. 퇴적률 상승과 식생의 증가가 반복되는 되먹임의 순환 속에서 식물과 지형은 어느 한쪽이 다른 쪽을 통제하는 일방적 관계보다는 서로가 영향을 주고받는 양방향의 능동적 상호관계를 형성한다(Stallins and Corenblit, 2018).
2) 생물지형학의 미래: 진화생물학을 포용하는 가설의 제안
Kim et al.(2024)은 최근 학계에 세 가지 종류의 선택압(selection pressure)을 포괄하는 개념적 모델을 제안하였다. 그림 2의 (a)에서는 전통적인 자연선택 개념에 의거하여 환경 조건이 직접적인 선택압이 된다. 생물종들 사이의 공진화(biological coevolution)를 표현하는 (b)에서는 환경 조건과 더불어 하나의 종이 다른 종(들)에게 미치는 직접적인 영향 역시 선택압이 된다(예: 식물과 곤충의 공생 관계). 마지막으로, (c)는 본 논문에서 새롭게 제안하는 생물지형학적 공진화 되먹임(biogeomorphological coevolutionary feedback)을 보여준다. 이 개념에서 각 생물종은 나름대로 자신의 서식 환경을 바꾸어 나간다. 그리고 이렇게 바뀐 서식처의 환경 조건은 자기 자신에게, 그리고 다른 종에게 새로운 선택압으로 작용할 수 있다. 생물지형학자들은 (c)를 다양한 종류의 선택압을 포용하는 통합적 시나리오로 간주하고, 이에 대한 장기간에 걸친 연구를 통해 기존의 자연선택 이론을 보다 포괄적으로 발전시킬 필요가 있다.
현실적으로 이 과업은 여전히 생태학자와 진화생물학자들에게 가설 수준에 머물러 있는 난제로서 신생・미개척 분야에 해당한다. 기존의 전통적인 다윈적 사고방식을 포용하되(Darwin, 1859; Dawkins, 1982), 도전적이고 혁신적인 접근 방법과 연구 질문을 집요하고 꾸준하게 개발・검증할 필요가 있다. 자연선택을 단어 그 자체로 해석한다면, 특정 환경 조건이 생존과 번식에 유리한 형질을 결정한다고 간단히 표현할 수 있다. 그렇다면, 생물종은 - 전술한 생물지형학적 관점에서 볼 때 - 자연선택의 과정에 능동적 참여자로서 개입할 수는 없을까? 위에 언급한 ‘특정 환경 조건’이 부분적으로라도 어떤 생물의 적극적인 생물지형학적 작용을 통해 형성되었음을 인정한다면(예: 비버의 둑), 그 생물종은 자신에게 가해지는 선택압을 - 의도하였건 아니건 - 일정 부분 스스로 만들어 내는 것은 아닐까? 그렇다면, 생물종은 자연선택의 과정과 속도에 어느 정도 영향을 줄 수 있지 않을까? 궁극적으로, 생물이 ‘자연선택을 선택’할 수 있다는 도전적이고 함축적인 표현을 우리 학계가 진지하게 고민하는 계기를 만들 수 있지 않을까?

그림 2.
선택압의 세 가지 종류: (a) 환경 조건, (b) 환경 조건과 생물종간 상호작용, (c) 환경 조건과 생물종간 상호작용, 그리고 생물에 의해 바뀌는 서식 환경(Kim et al.(2024)의 Fig. 1을 저자 재구성)
물론, 이와 비슷한 문제의식이 본 논문에서 최초로 제기되는 것은 아니다. 생태학과 진화생물학 분야에서는 각각 생태계 엔지니어링(ecosystem engineering)과 지위구축(niche construction)이라는 개념이 대두된 바 있다. 생태계 엔지니어링은 생물이 서식처의 생물적・환경적 조건을 바꿈으로써 다른 생물에게 주어지는 자원의 가용성을 직간접적으로 조절하는 현상을 뜻한다(Hastings et al., 2007; Jones et al., 1994, 2010). 지위구축은 생태계 엔지니어종이 스스로의 생존 가능성을 높이는 과정에서 신진대사와 활동, 선택 등을 통해 자신과 다른 종의 지위를 만들어 내거나 바꾸는 과정으로 정의된다(Odling-Smee, 1988; Odling-Smee et al., 2003).
본 논문은 생태계 엔지니어링과 지위구축 개념을 다루는 기존의 연구 흐름에 두 가지 보완점을 제안한다(Kim and Lee, 2022). 첫째, 대부분의 연구가 단일 생물종이 야기하는 생물지형학적 영향에 초점을 맞추고 있는데, 현실적으로 다른 종과의 직간접적인 상호작용 없이 단독으로 존재하는 종은 없다. 그러므로 그림 2의 (c)와 같이 최소한 둘 이상의 생물종을 동시에 고려하여 종합적인 생물지형학적 공진화 모델을 개발・검증할 필요가 있다. 둘째, 다수의 전통적 접근에서는 지위구축 형질이 시간에 따라 변화하지 않음을 가정하였다(Dong, 2022; Dong et al., 2024). 그러나 자연선택을 통한 진화는 특정 형질의 변화를 포함하기 때문에, 상황에 따라 유연하게 변모하는 지위구축 형질을 구현할 필요가 있다.
본 논문은 기존의 생물지형학적 연구의 흐름에 진화생물학적 관점을 명시적으로 접목하여 다음과 같은 세 가지 가설을 제안한다. 첫째, 둘 이상 생물종의 지위구축 형질과 서식처 환경 조건은 공진화하며 궁극적으로 각 종의 적합도(fitness)를 향상시킨다. 이렇게 향상된 적합도는 기존의 형질을 유지・고양하여 공진화 되먹임을 강화할 것이다(그림 3). 물론, 이 가설은 환경 조건의 급격한 변화나 천적의 과도한 포식과 같이 특정 개체군의 존속에 심각한 문제를 유발하는 드물고 극단적인 상황을 배제한다. 예를 들어, 해안습지에서는 식물과 게가 대표적인 생태계 엔지니어종으로서 상호작용하며 지표 경관을 변화시킨다. 게는 굴삭(또는 생물교란, bioturbation)을 통해 습지 토양의 배수(drainage)와 산화환원전위(oxidation-reduction potential)를 향상함으로써 식물의 생장률을 높인다. 식물은 뿌리 조직을 통해 토양의 안정도를 높임으로써 게의 굴삭을 촉진하는 것으로 알려져 있다. 이러한 생물지형학적 활동을 유발하는 지위구축 형질은 두 생물종의 서식 조건은 물론 이들의 적합도와 생식을 유지・향상시킬 것이다.
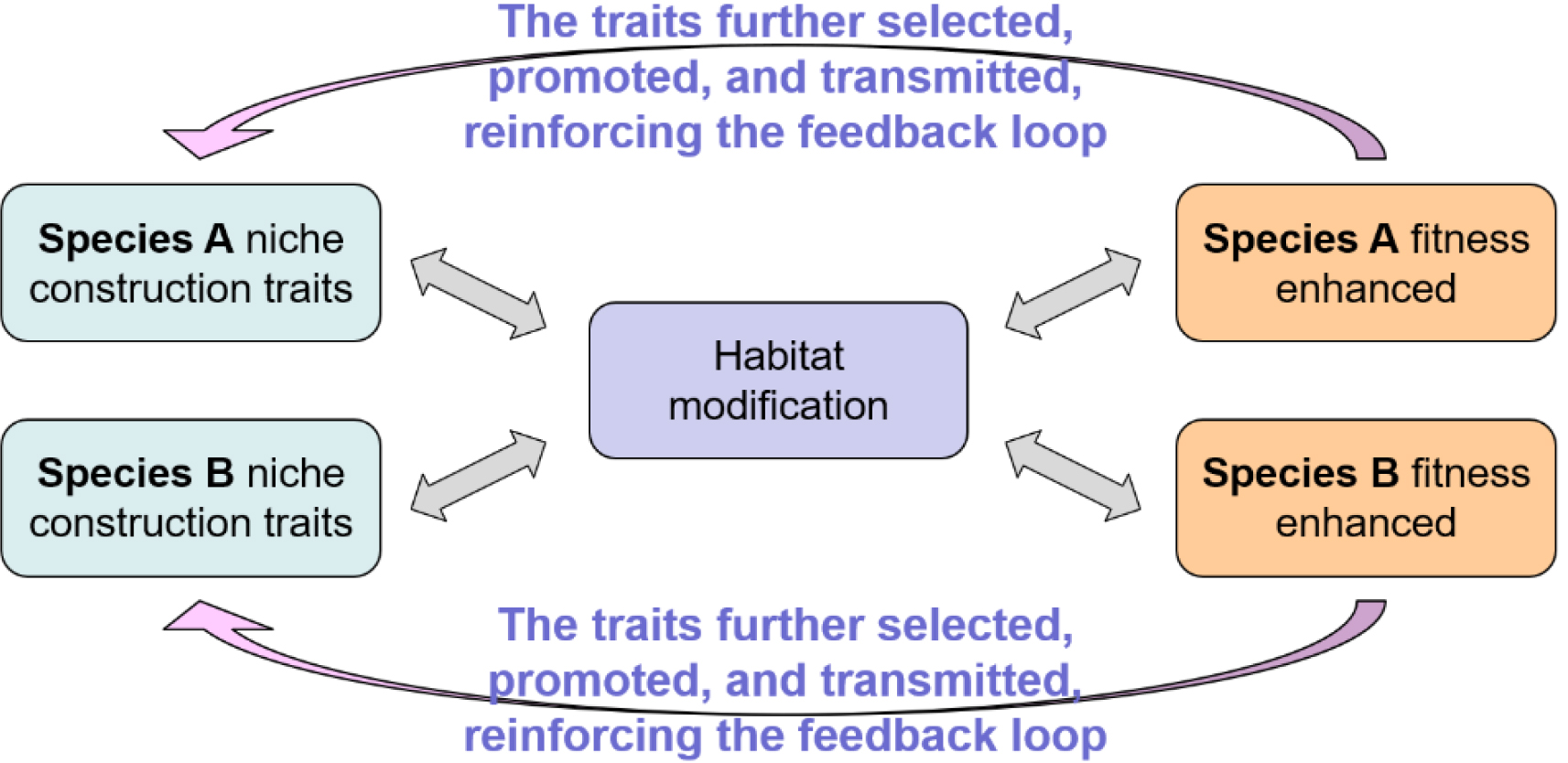
그림 3.
둘 이상의 생물종이 가지는 니치구축 형질과 이들에 의한 서식처 환경 조건 변화를 포괄하는 생물지형학적 공진화 되먹임 모델
(Kim et al.(2024)의 Fig. 7을 저자 재구성)
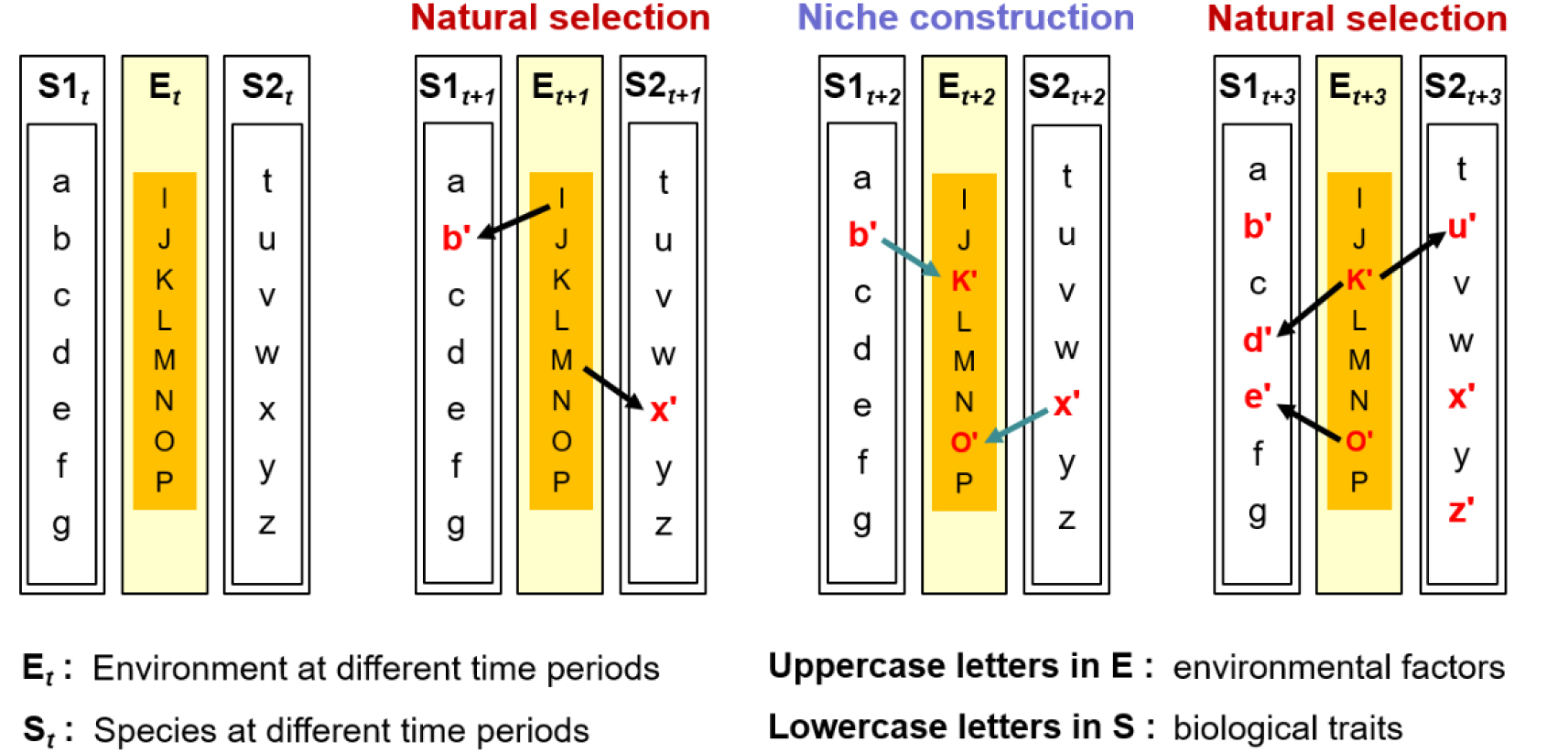
둘째, 생물종은 생물지형학적 활동을 통해 스스로의 서식처 환경 조건을 바꿈으로써 자연선택의 유형과 속도를 조절한다. 심지어, 억제할 수도 있다. 서식처 환경 조건이 그렇듯, 대부분 생물종의 지위구축 형질이 영원불변할 수는 없다. 해안습지의 사례로 돌아가서, 그림 4가 식물(S1)과 게(S2)뿐 아니라 환경(E)을 포괄하는 제4기 생물지형학적 공진화 모델을 구체화한다고 가정해보자. 각각은 알파벳 소문자 또는 대문자로 표현된 형질 또는 인자들로 구성되어 있다. 특정 시기 t에 간빙기가 도래하여 기온과 해수면이 모두 높아졌다. 이에 따라, 두 생물종은 향후 수만에서 수십만 년 정도는 바뀐 환경 조건, 특히 잦아진 침수로 인한 새로운 선택압에 대응할 필요가 생겼다. 침수는 식물에게는 혐기 조건(I)으로서 스트레스가 된다. 전통적인 진화생물학의 관점에서는 증가한 혐기 조건을 견디는 생리적 형질을 갖춘 개체가 자연선택의 유력한 대상일 것이다. 그러나, 본 논문에서 추구하는 생물지형학적 관점에서는, 퇴적률을 극대화하여 습지 고도를 높임으로써 자신에게 가해지는 혐기 스트레스를 줄이는 능력을 갖춘 개체 역시 자연선택의 대상이 될 수 있다. 즉, 조류를 감소시키고 부유 물질을 포집하는 기존의 형질(b)이 보다 두드러진 형질(t+1 시점의 b')로 자연선택이 발생하는 것이다. 이 형질을 갖춘 개체들이 우점하는 상황에서는 활발한 퇴적 작용을 통해 습지 고도가 꾸준하게 상승하고 다량의 유기물을 함유한 퇴적층이 형성되는 지위구축 효과가 기대된다(t+1의 K → t+2의 K'). 한편, 게로서는 t 시점에 증가한 침수로 인해 서식처의 토양수분 함량이 높아져(M) 굴삭을 통해 마련해 놓은 땅속 터널이 붕괴될 가능성이 높아졌다. 자신의 분비물과 집게를 사용하여 터널 표면의 퇴적물 입자를 견고하게 다지는 기존의 형질(x)이 보다 두드러질 필요가 생긴 것이다(t+1 시점의 x'). 이 형질을 갖춘 개체들이 우점하는 습지에서는 토양의 배수와 통기(aeration), 산화환원전위 등이 향상되는 지위구축 효과가 예상된다(t+1의 O → t+2의 O'). 마지막으로, 식물과 게의 지위구축을 통해 변경된 서식처 환경 조건은 더 나아가 두 생물종에게 새로운 선택압으로 작용할 수 있다. 서식 조건 K'와 O'에서는 스트레스의 감소로 인해 식생의 밀도가 높게 유지되기 때문에 식물은 크게 자라고(t+3의 d') 깊은 뿌리 조직을 형성해야(t+3의 e') 경쟁 우위를 점할 수 있다. 게의 경우, 식물의 촘촘한 뿌리 조직을 극복하고 굴삭을 수행할 만큼의 강한 집게(t+3의 u')를 갖추어야 할 것이다.
셋째, 생물종은 스스로의 생물엔지니어링(bioengineering) 활동을 통해, 서식처의 안정성(stability)과 회복탄력성에 영향을 준다. 지난 30여 년간, 생태학과 진화생물학 분야에서는 생태계 엔지니어링과 지위구축을 통해 변화하는 생물종 사이의 관계와 진화 과정에 대해 많은 연구가 이루어졌다(Laland et al., 1999; Lewontin, 1983; Odling-Smee et al., 2003). 반면, 이러한 생물지형학적 활동이 다양한 시공간적 스케일에서 서식처 또는 전반적인 생태계의 안정성에 기여하는 바에 대해서는 알려진 바가 많지 않다(Vahsen et al., 2023). 전통적으로, 생물은 암석이나 토양 표면에 보호막을 형성함으로써 침식을 완화하고 퇴적률을 높이는 인자로 인식되어 왔다(김대현, 2021). 산사태 발생지나 도로 공사로 인한 절개지에 사방공사를 하며 식생 피복을 설치하는 것이 좋은 예가 된다. 생물지형학에서는 이를 흔히 생물의 보호작용(bioprotection)으로 부른다(Carter and Viles, 2005; Coombes et al., 2013). 본 논문은 생물에 의한 침식완화 효과에 시공간적으로 한계가 있음이 향후 연구에서 검증될 필요가 있다고 제안한다. 앞서 해안습지 사례에서, 식물과 지형 사이의 되먹임을 통해 습지 표면의 퇴적물은 뿌리 주변에서 안정적이며 고도는 점진적으로 상승함을 설명한 바 있다. 그러나 사실 이는 식생이 존재하는 장소에 해당하는 시나리오이다. 넓은 공간적 스케일에서 보면, 식생 피복이 완화하는 조류 에너지는 - 식물이 습지 전역을 균일하게 덮지 않는 한 - 필연적으로 주변의 나지나 식피밀도가 낮은 곳에 집중되어 그곳의 침식을 가속할 것이다(그림 5의 오른쪽; 김대현, 2021; Kim and Lee, 2022; van Wesenbeeck et al., 2008). 한편, 지형학적 측면에서 고도와 경사는 일반적으로 상당히 높은 양의 상관관계를 가지며(Kim et al., 2008, 2016b), 이는 해안습지 생태계에서도 마찬가지다. 식물로 인해 퇴적률과 고도가 상승한다면, 습지 표면은 장기적으로 가파른 형태를 띠게 된다(van de Koppel et al., 2005). 아울러, 식물 뿌리의 생물지형학적 작용을 통해 퇴적물 입자들은 강한 응집력을 가지게 된다. 이러한 조건으로 인해, 해당 습지 생태계는 높은 안정성과 회복탄력성을 유지하리라 기대되지만, 이는 수십에서 수백 년에 달하는 비교적 단기간에 걸친 예측일 뿐이다. 장기간에 걸친 고도의 상승은 필연적으로 경사의 상승을 야기하여, 높은 에너지의 파랑 발생시 입자의 응집력이 높은 습지 전면의 붕괴로 이어질 수 있다(그림 5의 왼쪽; van de Koppel et al., 2005). 종합하면, 그림 5에 묘사된 두 상황은 애초에 습지 전체적으로 식물이 존재하지 않았을 경우보다 식물이 존재함으로 인해 더 높은 침식이 발생할 수 있음을 보여준다. 이러한 생물에 의한 침식, 즉 생물침식(bioerosion) 개념은 지금까지 생물지형학계 내에서 많은 주목을 받지 못해왔다(김대현, 2021).
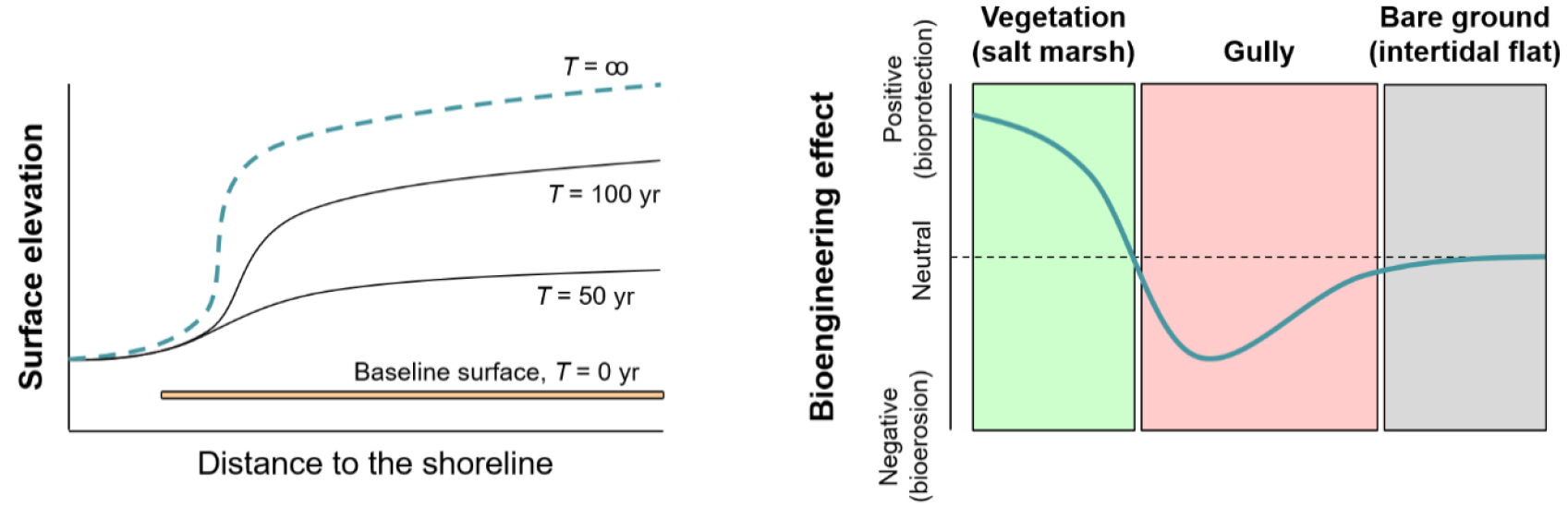
그림 5.
해안습지에서 생물지형학적 공진화 되먹임의 시간적 의존성
(좌; van de Koppel et al.(2005)의 Fig. 2를 저자 재구성)과 공간적 의존성(우; van Wesenbeeck et al.(2008)의 Fig. 2를 저자 재구성)
3) 소결
현재 우리나라 자연지리학계에는 생물, 수문, 지형, 토양 등 다양한 자연 요소를 아우르는 총체적인 연구의 틀은 물론, 이들 사이의 되먹임 관계를 공진화적으로 풀어가는 연구가 전무하다. 물론 각 요소에 대한 깊은 이해를 추구하는 전통적인 학문 분야는 충분히 존재하며 눈부신 이론적・기술적 발전을 이루어 왔다. 그러나 이렇게 생산되는 지식이 서로 단절되어 환원주의적 성격을 띠는 경향이 다분한 것도 사실이다. 이러한 한계를 극복하고 여러 학문간 통합을 추구하는 생물지형학은 지형학과 생태학, 진화생물학 같은 전통적인 학문 분류에 국한되지 않고 새로운 기초 학문 분야로 발전할 가능성이 충분하다. 이를 통해, 궁극적으로 우리나라 기초과학의 다양성 유지와 제학문간 융합을 달성하는 데 도움이 될 것으로 기대된다. 또한 본 논문에서 제안하는 생물지형학의 향후 발전을 통해 궁극적으로 자연선택 개념에 도전할 가능성이 열릴 것으로 여겨진다. 이 개념은 과거 자연선택설로 불리기도 하였으나, 오늘날 과학계에서 가설 수준을 넘어 널리 인정받는 이론으로 여겨지고 있다(최재천, 2009). 그러므로 본 논문에서 말하는 도전은 자연선택을 부정하거나 수정하는 의도를 가지지 않는다. 오히려, 이제껏 주목받지 못한 새로운 관점을 추가함으로써 이 개념을 보완하고 더욱 포괄적으로 발전시키는 과정을 의미한다.
5. 요약 및 결론
전통적으로, 시대상 무르익은 발견은 여러 분야의 과학자들이 독립적으로 작업하면서 이루어졌고, 이들은 당시 실현된 것 이상의 더 큰 발견을 예상하고 있었다(Whittaker, 1967). 역사상 한 획을 그은 개념이나 패러다임은 어느 한순간 단 한 명이 밝혀낸 것이 아닌, 약속이나 한 듯 비슷한 생각을 가진 동시대 학자들의 집단 지성이 모여 수렴된 것이다. 이런 인식을 바탕으로, 본 논문에서는 자연지리학과 생물지형학의 현재와 미래를 관통하는 시사점을 인지하고 관련된 혜안을 얻기 위해 지형학, 토양학, 생태학 분야의 과거 고전 논문을 검토하였다. 이를 통해, 두 학문 분야의 밀접한 관계를 확인하였고, 학제 간 연결이 생물지형학 분야로 수렴하는 과정에서 새로운 통찰력을 얻을 수 있었다.
이러한 논의는 결국 우리나라 자연지리학의 과거와 현재를 조망하고 앞으로 새롭게 나아갈 길을 모색한다는 본 논문의 궁극적인 목적과 맞닿아 있다. 자연지리학은 기권, 수권, 암석권, 생물권을 아우르고, 기상・기후학, 수문학, 지형・지질학, 토양학, 생물・생태학 분야의 다양한 학문적 원리를 바탕으로 하는 총체적인 시공간 과학이다. 그러나 현실적으로 우리나라 자연지리학의 학문사적 흐름에서 생물권에 대한 관심과 연구가 크게 부족했음은 부인할 수 없는 사실이다(김대현, 2021). 이런 상황은 자연지리학 가치와 지평을 스스로 제한하는 결과를 가져올 수 있다. 복합학문으로서의 다양성과 총체성을 추구하기 어려운 점은 물론, 생물학, 생태학, 생태공학을 비롯한 타 학문과의 교류에 제한이 생길 수도 있다. 무엇보다도 자연지리학에 매진하는 학문후속세대가 환경 및 종다양성 문제 해결에 기여할 여지가 줄어들 수 있음을 간과할 수 없다.
관현악의 본질이 여러 악기의 상호작용과 조화가 주는 종합적 감동에 있듯, 자연지리학자가 생산하는 지식은 지구 시스템을 구성하는 4대 권역을 개별적으로 연구하는 기존 자연과학 분야의 산술적인 덧셈 이상을 의미한다. 열린 시스템의 구성 요소간 상호작용과 되먹임, 그리고 이로부터 파생되는 비선형성을 다루는 생물지형학은 향후 자연지리학의 정수가 될 것으로 기대된다. 사실, 생물지형학을 지형학, 생태학, 진화생물학을 포괄하는 총체적 과학으로 정의한 김대현(2021)의 관점에서는 국제 학계로 시선을 돌리더라도 생물지형학이 여전히 태동기를 벗어나지 못하였다고 여겨진다. 이 세 학문 분야를 융합하는 진정한 의미에서의 생물지형학은 전세계적으로도 여전히 극소수의 과학자들만이 시도중이다(Dong, 2022; Dong et al., 2024; Kim et al., 2024; Vahsen et al., 2023). 우리나라 지리학계에 생물지형학이 자리를 잡고 번성하는 데 초석을 다지는 노력이 지속적으로 필요하다. 여러 후속세대 학자들이 자연지리학적 배경을 바탕으로 진화생물학적 소양을 닦아 생물지형학이라는 국제적 틈새시장에서 활약하는 미래를 기원한다.
